ISSN : 0975-3710
EISSN : 0975-9107

GLYN HOBBS1*, ISMINI NAKOUTI2
1School of Pharmacy and Biomolecular Sciences, Liverpool John Moores University, Byrom Street, Liverpool, L3 3AF, UK.
2School of Pharmacy and Biomolecular Sciences, Liverpool John Moores University, Byrom Street, Liverpool, L3 3AF, UK.
* Corresponding Author : g.hobbs@livjm.ac.uk
Received : - Accepted : - Published : 09-04-2012
Volume : 4 Issue : 3 Pages : 202 - 206
Int J Agr Sci 4.3 (2012):202-206
DOI : http://dx.doi.org/10.9735/0975-3710.4.3.202-206
Conflict of Interest : None declared
Organisms were isolated on their basis of survival in an iron-limited environment. The survivors of this treatment were largely actinomycetes. Of the viable cultures, most were found to produce siderophore like compounds. The most prolific producers as assessed by the Chromo Azuerol Sulphate assay were further characterised and found to belong to the genus Streptomyces. Attempts to taxonomically characterise these organisms illustrated the conserved nature of the 16S rRNA gene in this group of organisms. Physiological characterisation was undertaken and it was found that the resistance of these organisms to a range of antibiotics proved to be a useful discriminating factor .
Actinomycetes, iron chelation, siderophores, soil, streptomycetes, classification, resistance
Iron is a fundamental element to microbial nature. Due to the evolution of the di-oxygen atmosphere in the earth’s terrestrial environment, iron forms insoluble, bio unavailable ferric moieties at physiological pH [1] In order to capture iron microorganisms have developed specific mechanisms involving the formation of siderophores [2] The latter are low molecular weight compounds synthesised under iron starvation; they are produced extracellularly to bind iron and other ions forming a metal-siderophore complex. These complexes are then bound by the cells and transported intracellularly [3-6]
The actinomycetes are notorious environmental scavengers and have well-developed processes to capture ions in conditions depleted of elements; they are therefore an ideal target group for exploitation in ion binding technology [7-9] Here we report the characterisation of four siderophore producing actinomycetes.
The taxonomy of streptomycetes has been extensively studied by many groups and comprehensive reviews are available in the literature [10-12] Modern molecular taxonomical approaches involve the sequence analysis of the 16S rRNA gene, a particularly powerful tool in the classification of streptomycetes. Although 16S rRNA is a highly conserved area of the genome, it contains enough variations to allow us to investigate relationships at the genus, species and strain levels. Nevertheless it can be misleading due to intragenic variation, which highlights the need of genotypic and phenotypic observations [12] One possible discriminatory factor is the resistance profile to antibiotics. This article describes data that uses antibiotic resistance to differentiate the siderophore producing isolates.
A soil sample (1g) was suspended in 100ml of sterile distilled water and incubated 400 C for 24 hours. Actinomycetes were isolated from the soil suspension using a dilution plate technique on starch-casein agar (1.0 of starch, 0.4g of casein, 0.5 g of KNO3, 0.1 g of MgPO4, 0.2 g of K2HPO4, 0.1 g of CaCO3 and 15g of agar per litre of distilled water [dH2O] supplemented with 150 mM 2,2’-dipyridyl (DIP). The agar plates were incubated at 270 C for up to four weeks. Selective colonies were further sub-cultured and pure actinomycetes were isolated [13]
Siderophore production was confirmed using the Chromo Azuerol Sulphate (CAS) assay (Renshaw et al. 2003). Arnow’s [14] and Atkin’s [15] assays confirmed the presence of a catecholic and hydroxamate compounds respectively.
Actinomycete colonies were characterised according to Bergey’s Manual of Systematic Bacteriology [16] specific characteristics were observed by light and scanning electron microscopy.
The ability of the organisms to utilise different nutrient sources was tested using the SF-N2 (OXOID) and GP2 (OXOID) Biolog Microplates.
Antibiotic resistance of the strains was examined using MASTRINGS (Mast Diagnostics) and susceptibility discs (OXOID) performing the disc diffusion test. The model streptomycete, Streptomyces coelicolor (S. coelicolor) was also included in this study for comparison.
The strains were also tested for their ability to grow and sporulate on a range of liquid or solid media, including yeast extract-malt extract (YEME), Mannitol-Soya bean flower (MS), Muller-Hinton (MH), MH glucose, MH fructose and starch-casein agar [13]
Genomic DNA was extracted using the ‘Kirby mix procedure’ [13] Amplification of the 16S sequences were performed according to Rintala et al. 2001 [17] amplified products were purified using the QIAquickR PCR purification kit (50) [Qiagen] and sent to LarkTM Technologies, Inc (United Kingdom) according to their requirements.
Pairwise sequence comparisons and retrieval of homologous sequences were conducted by the NCBI BLAST database (available online- http://www.ncbi.nih.gov/). Similar sequences, including an outgroup (ancestral sequence), were multiple aligned by BioEdit sequence alignment Editor 7.0.4.1 (Copyright 1997-2005, Hall, 1999). Taxonomic analysis was performed by the ClustalX software, version 1.83 and Bootstrap confidence values (1000) were provided by the Neighbour- Joining (NJ) algorithm. Finally the phylogenetic tree was presented using TreeView (Win 32) 1.6.6 (Copyright 2001) by Roderic. D. M. Page, available online: http://taxonomy.zoology.gla.ac.uk/rod/rod.html).
In an attempt to identify novel siderophore compounds we have isolated and purified 154 cultures from soil samples collected in Thailand (Tak province). The majority of strains are considered to be members of actinomycetes on the evidence of phenotypic examination. All the isolates were grown on starch-casein agar containing 150 mM DIP. This concentration of DIP was chosen as it reduced the number of surviving actinomycete colonies by approximately 99%. The remaining 1% was thought to be likely to have physiological attributes that enabled them to survive iron limitation.
Twenty-two strains were confirmed CAS-assay positive with five strains (23F, 31B, 31C, 33D and 29C- GenBank accession numbers EF585403 - EF585407) exhibiting prolific siderophore production. Liquid based assays were conducted on culture supernatants in order to categorise the siderophore producers. The results classified the strains 31B, 31C, 33D and 29C as catechol producers, whereas 23F was categorised as a hydroxamate synthesiser.
16S rRNA gene sequence analysis was carried out to elucidate the taxonomic position of the isolates. Almost complete 16S rRNA sequences of the representatives were determined following the isolation of genomic DNA and sequencing of the amplified genes. The strains’ phylogenetic positions are reported in [Fig-1] . The levels of 16S rRNA sequence similarities between the isolates and actinomycetes species ranged from 97-99 %, confirming that the strains are members of the family, mainly streptomycetes.
The high percentage similarity values amongst species of the same genus are very common due to the highly conserved nature of the gene. For example although Streptomyces macrosporus and Streptomyces megasporus share 98 % similarities, they are clearly different on their phenotypic characterisation. A second paradigm includes Streptomyces thermocarboxydovorans (S. thermocarboxydovorans) and Streptomyces thermodiastaticus (99 %) or S. thermocarboxydovorans and S. thermoviolaceous (98 %) [18] Moreover Amycolatopsis albidoflavus share 97.4 % similarities with Amycolatopsis azurea and Amycolatopsis orientalis [19]
All the streptomycete stains were further investigated in order to determine their morphological, physiological, growth and biochemical characteristics. Their ability to use a variety of nutrient compounds is described in [Table-1] .
Table 1- Nutrient utilisation of strains 29C (GenBank accession number EF585407), 33D (GenBank accession number EF585406), 31B (GenBank accession number EF585404), 31C (GenBank accession number EF585405) and 23F (GenBank accession number EF585403).
Strain 29C is a Gram-positive, aerobic, white mycelia-forming organism that undergoes the typical streptomycete life cycle resulting in pink spirales spores. The spore surface was found to be smooth and this stage of the life cycle was only ever observed on starch-casein and YEME solid media. It did not sporulate on MS (Manitol-Soya agar) agar but it produced a diffusible pigment. It tolerated 150mM DIP and in starch-casein media it overproduced a very ‘powerful’ catechol that formed strong ferric complexes. It was found to produce a bactericidal agent against Bacillus subtilis (B. subtilis) and Candida albicans (C. albicans) under all conditions tested. In our hands it grew best in starch-casein media, pH 7.5 at 300 C.
It is a Gram positive, aerobic streptomycete species producing brown spirales spores. It grew rapidly on MS, starch-casein, yeast extract-malt extract (YEME) and slightly slower on Muller-Hinton-Fructose (MHF) or Muller-Hinton-Glucose (MHG). It was highly pigmented when grown on YEME and MS; nevertheless it did not sporulate well in the former cultivation media.
It grew poorly on MS, pH 5, and appeared to be unstable at pH 7. Optimum growth was on MS, pH 8 at 300 C. It produced an antimicrobial against B. subtilis and was found to be extremely tolerant to DIP (600mM), suggesting a well-adapted organism in its environment. It is worth noting that it overproduced a catechol on starch-casein agar, whereas it synthesised only a small amount of the specific siderophore on MS.
31C is a Gram positive, aerobic streptomycete producing rectiflexible white/grey spore chains. This strain only produced substrate mycelia on MHG or MHF agar, and on YEME solid media appeared to be extremely unstable forming heterologous colonies. It grew on MS agar, pH 5-7, with a preference for pH 7 where it only formed aerial mycelia. It solely sporulated on starch-casein media with an optimum growth at pH 7.5 at 300 C. It posed antibiosis against both B. subtilis and C. albicans but only in media where catechol production is negligible.
This isolate is a Gram positive, aerobic streptomycete producing rectiflexible spores. It was apparent that 31B had fastidious growth requirements and appeared unstable in our hands. It was also evident that its growth rate was extremely slow with a preference to starch-casein media, pH 7.5, at 300 C.
This strain is a Gram positive, aerobic actinomycete, which belongs to Streptomyces genus, and produced spores with an ornamented surface (spiny).
The organism was well adapted to its environment tolerating up to 600 mM DIP and growing in a range of pH (5-8) and temperature (280 C- 370 C) conditions. It grew quickly on MS (3 days) agar, nevertheless much slower in starch-casein medium (approximately 10 days). It did not sporulate on YEME agar and on MHG and its optimum growth conditions involved cultivation on MS or starch-casein, pH 7, at 370C.
Antibiotic resistance data offer valuable information in terms of strain characterisation and identification [Table-2] . In the specific small-scale experiment 14 out of 24 antibiotics demonstrated different patterns amongst the streptomycetes. Careful examination of the data revealed specific profiles for Streptomyces coelicolor (S. coelicolor) and strains 23F, 29C and 31B. For example sensitivity to amoxicillin, cefuroxim or oxaxillin was a unique characteristic of isolate 29C. Stain 31B was the only candidate that was resistant to vancomycin. A combination of growth in the presence of erythromycin and levofloxacin differentiated strain 23F from S. coelicolor. Culture 33D could be identifiable by its sensitivity against ampicillin and vancomycin. Finally S. coelicolor demonstrated a distinctive pattern of tetracycline (10 μg) and erythromycin resistance.
It is anticipated that the isolation and characterisation of new actinomycetes might lead to novel siderophores of significant pharmaceutical interest. Although 16S rRNA analysis is a very powerful tool in phylogenetic relationships, it does not provide enough evidence for classification of closely related species. Therefore a ‘universal’ approach, which will include the combination of modern molecular approaches with a number of morphological, physiological and biochemical characteristics, will facilitate the understanding of the genotypic and phenotypic behaviour of these precocious siderophore producers. Antibiotic resistance data provide useful information on the nature of the cells. A report by Hopwood reaffirms the importance of actinomycetes as a major reservoir of antibiotic resistance genes [20] Clearly these characteristics can be a useful way of classifying strains. This could be a valuable evolutionary route to differentiate strains within a population. Given the fact that Streptomyces species have been found to have the capacity to produce multiple “antimicrobial†compounds [] 21] and hence the ability to be resistant to these compounds, it will be interesting to develop a resistance fingerprint system for strains and to revisit phylogenetic analysis in the light of this information.
[1] Wandersman C. and Delepelaire P. (2004) Annu Rev Microbiol 58, 611-647.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[2] Gunter K., Toupet C. and Schupp T. (1993) J Bacteriol 175, 3295-3302.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[3] Neilands J.B. (1995) J Biol Chem 270, 26723-26726.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[4] Kalinowski B.E., Liermann L.J., Givens S. and Brantley S.L. (2000) Chem Geol 169, 357-370.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[5] John S.G., Christy E., Ruggiero L.E., Tung S. and Neu M.P. (2001) Environ Sci Techol 35, 2942-2948.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[6] Rodriquez M.G. and Smith I. (2003) Molecul Microbiol 47, 1485-1495.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[7] Meiwes J., Fiedler H.P., zahner H., Konetschny-Rapp S. and Jung G. (1990) Appl Microbiol Biotechnol 32, 505-510.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[8] Challis G.L. and Ravel J. (2000) FEMS Microbiol lett 187, 111-114.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[9] Bentley S.D., Chater K.F., Cerdeno-Tarraga A.M., Challis G.L., Thomson N.R., James K.D., Harris D.E., Quail M.A., Kieser H., Harper D., Bateman A., Brown S., Chandra G., Chen C.W., Colllins M., Cronin A., Fraser A., Goble A., Hilalgo J., Hornsby T., Howarth S., Huang C.H., Kieser T., Larke L., Murphy L., Oliver K., O’Neil S., Robbinowitsch E., Rajandream M.A., Rutherford B.G., Rutter S., Seeger K., Saunders D., Sharp S., Squares S., Taylor K., Warren T., Wietzorrek A., Woodward J., Barrell B.G., Parkhill J. and Hopwood D.A. (2002) Nature 417, 141-147.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[10] Williams S.T., Goodfellow M., Alderson G., Wellington E.M.H., Sneath -P.H.A. and Sackin M.J. (1983) J Gen Microbiol 129, 1743-1813.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[11] Williams S.T., Goodfellow M., Wellington E.M.H., Vickers J.C., Alderson G., Sneath P.H.A, Sackin M.J. and Mortimer A.M. (1983) J Gen Microbiol 129, 1815-1830.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[12] Anderson A.S. and Wellington E.M.H. (2001) Int J Syst Evol Microbiol 51, 787-814.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[13] Kieser T., Bibb M.J., Buttner M.J., Chater K.F. and Hopwood D.A. (2000) Streptomyces genetics, 8, 168-169 & 342-343. The John Innes Foundation, Norwich.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[14] Arnow L.E. (1937) J Biol Chem 118, 531-537.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[15] Atkin C.L., Neilands J.B. and Phaff H.J. (1970) J Bacteriol 103, 722-733.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[16] Locci R. (1989) Streptomyces and related Genera. In Bergey’s Manual of Systematic Bacteriology ed Williams Wikins Company, Baltimore. 2451-1508.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[17] Rintala H., Nevalaines A., Ronka E. and Suutari M. (2001) Mol Cell Probes 15, 337-347.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[18] Kim C., Walter Lorenz W., Hoopes J.T. and Dean F.F.D. (2001) J Bacteriol 183, 4866-4875.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[19] Lee S.D. and Hah Y.C. (2001) Int J Syst Evol Microbiol 51, 645-650.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[20] Hopwood D.A. (2007) Mol Microbiol 63, 937-940.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[21] Omura S., Ikeda H., Ishikawa J., Hanamoto A., Takahashi C., Shinose M., Takahashi Y., Horikawa H., Nakazawa H., Osonoe T., Kikuchi H., Shiba T., Sakaki Y. and Hattori M. (2001) Proc Natl Acad Sci USA 98, 12215-12220.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
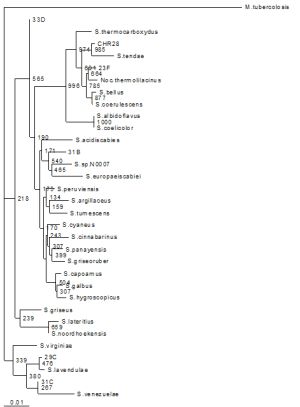 | Fig. 1- Phylogenetic relationships of the isolates and related Actinomycetes based on the 16S rRNA sequence analysis. The tree is bootstrapped by 1000 times by Neighbour-Joining (N-J). M. tuberculosis was employed as an outgroup |
 | Table 1- Nutrient utilisation of the five isolates |
 | Table 2-Antibiograms of the siderophore producing strains. S. coelicolor is included as a reference |