ISSN : 0975-5276
EISSN : 0975-9174

GAD EL-RAB D.A.1*, OLFAT S. BARAKAT2, IBRAHIM G.A.3, TAWFIK N. F.4, EL-KHOLY W.I.5
1Dairy Science Department, National Research Centre, Dokki, Cairo, Egypt
2Department of Microbiology, Faculty of Agriculture, Cairo University, Cairo, Egypt
3Dairy Science Department, National Research Centre, Dokki, Cairo, Egypt
4Dairy Science Department, National Research Centre, Dokki, Cairo, Egypt
5Dairy Science Department, National Research Centre, Dokki, Cairo, Egypt
* Corresponding Author : deiaa73@yahoo.com
Received : 17-04-2011 Accepted : 28-04-2011 Published : 15-07-2011
Volume : 3 Issue : 1 Pages : 59 - 66
Int J Microbiol Res 3.1 (2011):59-66
DOI : http://dx.doi.org/10.9735/0975-5276.3.1.59-66
Five Samples of traditional dairy products were collected from different rural areas in the Farafra region in west desert of Egypt. Thirteen isolates from these products were identified by Apparatus and Procedure of Identification (API) system; isolates were studied for their genetic diversity using PCR and their technological characteristics.
lactobacillus, Domiati cheese, Starters.
Lactic acid bacteria “LAB “are widely distributed in the nature. The lactic acid fermentation, which these bacteria perform has long been known and applied by the humans for making different foodstuff. It plays an essential role in the production of all dairy products and is involved in the production of many other foods and drinks. In recent years, probiotic activity of lactic acid bacteria has been emphasized. Probiotic is commonly defined as viable microorganisms that exhibited a beneficial effect on the health of the host when they are ingested. [18,10] , the most widely used probiotic bacteria are lactobacilli and bifidobacteria which can survive in the intestine. In the rural areas in Egypt, the wild LAB microbiota represents a natural reservoir for microbial cultures that contains diverse genetic information. It also has characteristic strains that are highly resistant to the manufacturing environmental conditions such as high salt concentration and phage infection. Moreover, they contribute to produce specific characteristics such as taste, aroma and texture of traditional Egyptian dairy products. However, modern large-scale production techniques generally make use of starter systems with defined strains so as to guarantee uniformity, safety and quality in the final product [23,20] . In our country at domestic conditions dairy products are prepared from cow, but also sheepish, goats and buffalo-cow’s milk. By these products the species composition of lactic acid bacteria is more various and inconstant when compared with those of the trade products. The wild strains in the biotechnological aspects are perspective as bacteriocin produces and probiotics. They have a potential use for the establishing of new the so called functional foods [14,17] . The aim of the recent study was the isolation and taxonomic determination of lactic acid bacteria from cheese (wild type) in order for their including in original Egyptian collection of lactic acid starters.
• Five samples of pickled white cheese were used as source to isolate lactic acid bacteria. They were gathering from Farafra Oasis in Egypt (which the weather is so hot up 400C).
• MRS broth which 1.6 %(w.v) of agar and 0.8 %(w.v) CaCo3 were added to isolate lactic acid bacteria strains. 25 gram of each sample was homogenized with 225 ml of 0.85% sterilized NaCl solution by shaking for several minutes. From the suspensions, serial dilutions were made in NaCl and plated by spreading 0.1 ml onto the surface of MRS agar. After anaerobic incubation at 300C for 48h colonies which dissolved CaCo3 and formed clear zones around their own colonies on the medium plate were isolate at random. The broths inoculated with each colony were cultivated at 300C and tested for catalase reaction. Isolated strains were stocked as frozen cultures in MRS broth with 15 %(w.v) glycerol at -800C.
• Gas production; Overnight cultures of LAB were inoculated into 10 mL MRS broth containing inverted glass tubes (approximately 6 × 50 mm) and incubated at 300C. Results were recorded after 48 h. Cultures producing gas within the 48-h incubation period were presumed to be heterofermentative LAB. Those cultures that did not produce gas within 48 h were presumed to be homofermentative LAB.
• Survival of the isolates in the presence of bile salts was determined by adjusting the early log phase MRS agar culture to pH 6 with 1 N NaOH and adding suspension of bile salts (Difco, Detroit, Michigan, USA) to obtain different concentration (0.0,0.1,0.2,0.3,0.4,0.5 %.) The viable bacterial counts were determined after 24 and 48h incubation at 370C.
• Antimicrobial activity Test to determine the inhibitory capacity of isolated LAB against each other’s and pathogenic strains such as Listeria monocytogenes 15313. The isolated LAB and pathogenic strains were incubated in MRS agar medium at 37°C for 24 to 48 h using Agar diffusion test.
• The isolates were also tested to produce exopolysaccharides (EPS) which form a slime layer loosely attached to the cell surface or secreted into MRS agar which able to produce strands when extended with an inoculation loops.
• Survival of the strains at low pH was determined by adjusting the early log phase at OD value ~ 0 .1 MRS cultures to pH 2 and 1.5 with 1 M HCl and incubation at 370C for 6 h. The viable bacterial counts were determined every 30 minute till 6 h on MRS agar. Respective survival % of cultures was calculated from the cell numbers.
• Isolates identified as the genus lactobacillus were subjected to carbohydrate fermentation. API 50 CH strips medium (BioMe´rieux, Marcy-l’Etoile, France). As recommended by the instructions. The corresponding APILAB Plus database and software were used for presumptive identification of species and reference to Bergey’s Manual of Systematic Bacteriology [30] .
• PCR (polymerase chain reaction) based method [1] was also conducted for specific lactobacilli isolates which were cultivated in MRS agar incubated under anaerobic conditions for 24 h at 370C. Isolates were identified to the genus level. The PCR products were analyzed by agarose gel electrophoresis and photographed under UV exposure. PCR products were purified using a QIAquick PCR purification kit (Qiagen, Inc., Valencia, CA), and the DNA fragments were sequenced. Sequences were analyzed by performing a BLAST (Basic Local Alignment Search Tool) search of the National Center for Biotechnology Information nonredundant DNA sequence database [http://www.ncbi.nlm.nih.gov/] .
• The specific growth rate “µ” was calculated from the slope of a linear regression line during exponential growth with a correlation coefficient (r2) of 0.99. Each point represents the mean of three independent cultures.
Formula 1 μ = (ln x – ln x0)/ (t – t0)
Where X is the optical density in the end and X0 in the beginning of the exponential growth phase and at time t and t0, respectively. Three independent replicate assays were performed, and the variations were less than 5%. Statistical analysis and graphical representations were performed using OriginLab Corporation software (Northampton, MA).
A total of thirteen lactic acid bacteria were isolated from the cheese [Table-1] , Isolates were gram positive and catalase negative they were also investigated to the ability of acid production in MRS broth after 24hrs of the growth.
Growth and survival of the thirteen isolates of Lactobacilli were comparatively studied; cultures were also identified to the level of species. However, for the strains used as an adjunct, the fast acid production may not be a desirable trait since it could lead to excessive acidification during cheese making.
None of the isolated strains produced gas in MRS broth 2% glucose at 28, 30, 37 or 45oC.
According to calculations (Formula 1), all isolates had significantly lower maximum specific growth rates compared to isolate number 401 (µ = 0.0.20, 0.28 and 0.13 h 1) and strain number 441 (µ = 0.14, 0.24 and 0.21 h 1) in MRS broth at 28,37, 45oC [Fig-1] . The highest growth rates of strain isolates were observed at 37oC then 45oC and after that by 280C which prove that kind of isolate are normally grow in desert weather where temperature usually high all the year.
The growth rates of isolate number 401 and isolate number 441 were in the different side, one of them prefer growing in low temperature and the other one prefer the high temperature which help to choose both of them to do extra research. As all strains grew well at 370C, this temperature was chosen for comparative growth experiments. Only two candidate strains (441 and 401) phenotypically selected for further study of their probiotic characteristics.
Antimicrobial activity was determined in direct antagonistic test; the strains belonging to the same species were tested against each other and Listeria monocytogenes 15313 in MRS agar plates. The results showed that of Lactobacillus isolates appeared to have no antimicrobial activity.
Many strains of LAB produce EPS that can be a capsule, closely attached to the bacterial cells or loosely attached or excreted as slime [2] . The strains that showed poor and good pellet separation were screened for EPS production. Approximately 12 strains of lactobacilli isolates were found to produce EPS slime. Among these strains, isolate number 441 was forming heavy exoploycharides (data not shown).
Isolates were also identified to the level of species by using fermentation profiles of selected Lactobacillus strains after anaerobic growth in MRS broth at 370C, Despite differences in sugar fermentation profiles of the strains comes from different groups, the initial API identification of strain 401 as Lactobacillus casei subsp. rhamnosus was confirmed by 16 r RNA gene sequencing on contrast the strain 441 the results of API was different from 16 r RNA gene sequencing analysis which clearly showed the correct affiliation of the strain to Lactobacillus paracasei subsp paracasei [Table-2] . Our results from PCR suggest satisfactory differentiation at the strain and species levels of the examined isolates.). Genetic taxonomy based on phylogenetic data, such as 16S rRNA, has shown the relatedness of the currently recognized and reorganized Lactobacillus species. As a consequence, some species previously included in the genus are now affiliated to a separate genus. Although phenotypic techniques such as the API system are still taken as powerful tools capable of discriminating between the species of Lactobacillus, the use of genetic methods for Lactobacillus taxonomy has become the backbone of reliable identification [25,28] .
Strains of 441 and 401 were most resistant to the salt, their growth was negligible at 10% NaCl as shown in [Fig-2] . The specific growth rates of strains 441 and 401 in the medium containing different concentration of salt 0 : 10% salt were (µ = 0.20 and 0.18 h 1), respectively at 1% salt and were (µ = 0.075 and 0.035 h 1), respectively at 10%salt which the most tolerant salt concentration could even grow in it. Salt addition influenced the morphology of cells. Generally longer or thicker cells and shorter chains were observed, in this respect [5] recorded that the growth rates of the L. paracasei and L. rhamnosus decreased 20–80% at salt concentrations 6–6.5%. The graph show ability of isolate 441 to grow better than 401 and this show how Egyptian dairy products are generally produced under artisan conditions using high salt concentrated solutions which allowed preservation for extended periods of their products which agree with [6,9] reported that strains of non-starter lactic acid bacteria (NSLAB) such as Lactobacillus paracasei can grow to 108 CFU/g during manufacture and ripening of cheddar cheese, highlighting the ability of NSLAB to grow in the presence of salt concentrations normally present in Cheddar cheese (~2.7%, w/w).
Stability of LAB to heating suggests that NSLAB in cheese may originate from the microflora of raw milk by surviving pasteurization. Both two tested isolates 441 and 401 were resistant to heating at 60oC for 30 min in MRS broth at pH 6 [Table-3] and [Table-4] , [Fig-3] . The isolate of 441 reduced by two logs while isolate 401 reduced by four logs after 15 min, in other hand the decimal reduction time D of isolate 441 was greater than isolate 401 which clear that 441 could tolerate heat more than 401. Survival of all L. paracasei strains during heating supports the idea that one source of NSLAB in cheese should be raw milk as also suggested by [16] . In contrast; non-starter lactococci studied by [7] did not survive at 600C for 30 min. It may be that some lactobacilli cannot be detected in milk immediately after heat treatment, but after a suitable recovery period they can be cultured which agree with our recovery results after three days, as they are heat-damaged but not actually killed. Obviously after a recovery period the numbers of cells may have been higher and the possibility exists.
In many microorganisms, exposure to sublethal pH induces an acid tolerance response (ATR) that significantly enhances cell survival under more extreme acid environment [24] . Induction of ATR improved survival (up to 100-fold) of isolates during acid challenge, The majority of the LAB possess an inducible acid tolerance response (ATR), also known as the acid adaptive response, which improves the survival of adapted cells upon exposure to lethal acid challenge [4] . Sensitivity to pH 2 or pH 1.5 (370C) of the strains studied was different and seems to be species specific [Fig-4] . Isolates of 441 and 401 keep viability at pH 2 or pH1.5 already during 6 h incubation. Survival of the strain 441 was lost two logs after 6 hours when challenge to acid condition pH 2 but was losing three logs in case of 401 in the same condition. At pH 1.5 a three log reduction was observed in strain 441, whereas a nearly four log reduction was observed in strain 401 after 6 hours of acid challenge which clear the strain 441 is could survive better than other strain 401, this may due to production polysaccharides which could reduce the lethal effect of low pH on the cell wall, The results demonstrated that 441 is significantly more resistant to acid than 401 isolate.
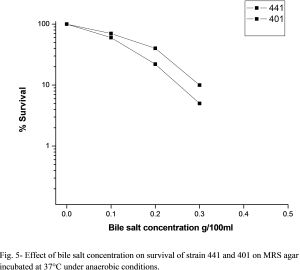
When evaluating the potential of using lactic acid bacteria as effective probiotics it is generally considered necessary to evaluate their ability to resist the effects of bile acids [18] . Oxgall is a natural dried bovine bile component containing both conjugated and unconjugated bile salts. Decrease of viable bacterial numbers (log cfu/ ml) with increase of presence of 0.0, 0.1, 0.2, 0.3, 0.4, .05% Oxgall g/100 ml MRS is shown in [Fig-5] . Survival was calculated from the actual numbers (cfu ml-1). The two bacterial strains were tested for can be viable till 0.3 % oxgall, after this concentration (in 0.4 and 0.5%) there is no viable cell could grow [21] .
The species of Lactobacillus growing in Egyptian traditional white cheese identified and classified by molecular methods could also be distinguished by physiological characteristics like maximum specific growth rates under different growth conditions, which can be established by simple cultivation experiments. The maximum specific growth rates of selected strains 401 and 441isolates were significantly different compared to other isolates in MRS broth at different tested temperatures. Growth of 441 and 401 were more resistance to salt and high acidity. Survived heating at 600C and tolerated bile and salt. Never theless, they may be of great interest for cheese production because of their ability to grow in ripening and high salted cheese. Generally, the viability is not affected when LAB are incubated with hydrochloric acid at pH 3.5 or higher [3,9] , while at lower pH the survival decreases to less than 1%, In addition, the strains 401 and 441 had the best survival capacities at pH (1.5 and 2), They survived under low pH conditions for 6 h and they tolerated well the low pH under in vitro conditions even at concentrations higher than those previously used by other authors [8,15] , Acid tolerance of bacteria is an important factor as well as to assure their resistance of gastric stresses also for their use as dietary adjuncts in acid foods.
[1] Altermann E., Russell W.M., Azcarate-Peril M.A., Barrangou R., Buck B.L.,McAuliffe O., Souther N., Dobsn A., Dobson A., Duong T., Callanan M., Lick S., Hamrick A., Cano R. and Klaenhammer T.R. (2005) Proc Natl Acad Sci USA. 102:3906–12.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[2] Cerning J. (1995) Lait, 75, 463–472.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[3] Chou L. S., and Weimer B. (1999) J. Dairy Sci. 82:23–31.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[4] Cotter P.D. and Hill C. (2003) Mol. Biol. Rev. 67:429–453.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[5] Crow V., Curry B., and Hayes M. (2001) International Dairy Journal, 11(4–7), 275–283.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[6] Desmond C., Stanton C., Fitzgerald G.F., Collins K. and Ross P.R. (2002) Int Dairy J; 12: 183-90.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[7] Durlu-Ozkaya F., Xanthopoulos V., Tunail N. and Litopoulou-Tzanetaki E. (2001) Journal of Applied Microbiology, 91, 861–870.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[8] Fernandez M.F., Boris S. and Barbes C. (2003) J Appl Microbiol, 94:449–455.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[9] Gardiner G., Ross R.P., Collins J.K., Fitzgerald G. and Stanton C. (1998) Appl Environ Microbiol, 64: 2192-9.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[10] Gill H.S. and Guarner F. (2004) Postgraduate Medical Journal, 80, 516–526.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[11] Holzapfel W.H., Haberer P., Geisen R., Bjorkroth J. and Schillinger U. (2001) American J. Clinic Nutr., 73: 365–373.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[12] Jacobsen C.N., Rosenfeldt Nielsen V., Hayford A.E., Møller P.L., Michaelsen K.F., Pærregaard A., Sandstrom B., Tvede M. and Jakobsen M. (1999) Appl Environ Microbiol, 65:4949–4956.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[13] Jordan K.N., and Cogan T.M. (1999) Letters of Applied Microbiology, 29 (2), 136–140.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[14] Kitazawa H., Ueha S., Itoh S., Watanabe H., Konno K., Kawai Y., Saito T., Itoh T. and Yamaguchi T. (2001) Int J Food Microbiol., 65, 149-162.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[15] Lee Y-K. and Salminen S. (1995) Trends Food Sci Technol, 6:241–5.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[16] Marteau P., Minekus M., Havenaar R. and Huis in’t Veld J.H.J. (1997) J. Dairy Sci. 80:1031–1037.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[17] Morelli L. (2007) International Dairy Journal, 17, 1278–1283.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[18] Noriega L., Cuevas I., Margolles A., De los R-G. and Clara G. (2006) Int Dairy J, 16:850–5.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[19] Oberman H. and Libudzisz Z. (1998) Fermented milks: microbiology of fermented foods. Blackie Academic Professional and London, 1:308–350.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[20] Ross R.P., Morgan S. and Hill D. (2002) Int. J. Food. Microbiol., 79, 3–16.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[21] Van de Guchte M., Serror P., Chervaux C., Smokvina T., Ehrlich S.D. and Maguin E. (2002) Antonie Van Leeuwenhoek, 82:187–216.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[22] Vandamme P., Pot B. and Gillis M. (1996) Microbiological Reviews, 60, 407-438.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[23] Ward D.M., Weller R. and Bateson M.M. (1990) Nature 345:63–65.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[24] Wijtzes T., de Wit J.C., Huis in’t Veld J.H.J., van’t Riet K. and Zwietering M.H. (1995) Applied and Environmental Microbiology, 61, 2533–2539.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[25] Wood B.J.B. and Holzapfel W.H. (1995) The Lactic Acid Bacteria: The Genera of Lactic Acid Bacteria, 1st edition. Chapman and Hall, London.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
 | Fig. 1- |
 | Fig. 2- |
 | Fig. 3- |
 | Fig. 4- |
 | Fig. 5- |
 | Table 1- Morphological properties of the isolated strains + Positive, - negative |
 | Table 2- The API 50 CHL System Kit and 16S rRNA sequencing results |
 | Table 3- Survival capacity (%) of strain 401 to thermisation (600C) from 0: 30 min) in MRS broth at pH 6.5 Strain 401 Lactobacillus rhamnosus subsp. rhamnosus, a = viable cell counts on MRS agar after thermisation, b = decimal reduction time, the time taken to reduce the cell numbers by one log cycle* Initial number for 401 log N = 11.04 |
 | Table 4- Survival capacity (%) of strain 441 to thermisation (600C) from 0: 30 min) in MRS broth at pH 6.5 Strain 441 Lactobacillus paracasei subsp paracasei, a = viable cell counts on MRS agar after thermisation, b = decimal reduction time, the time taken to reduce the cell numbers by one log cycle* Initial number for 441 log N = 11 |