ISSN : 0976-8823
EISSN : 0976-8831

SHEIKH S.A.1, WANI M.R.2, LONE M.A.3*, TAK M.A.4, MALLA N.A.5
1Department of Botany, Government Degree College (Boys), Anantnag-192 102, J&K, India.
2Department of Botany, Government Degree College (Boys), Anantnag-192 102, J&K, India.
3Department of Botany, Government Degree College (Boys), Anantnag-192 102, J&K, India.
4Department of Botany, Government Degree College (Boys), Anantnag-192 102, J&K, India.
5Department of Botany, Government Degree College (Boys), Anantnag-192 102, J&K, India.
* Corresponding Author : ahmadmansoor21@yahoo.com
Received : 07-09-2012 Accepted : 17-09-2012 Published : 20-09-2012
Volume : 2 Issue : 1 Pages : 34 - 38
J Plant Genom 2.1 (2012):34-38
DOI : http://dx.doi.org/10.9735/0976-8823.2.1.34-38
Sodium azide (SA) - a respiratory inhibitor has long been considered as a potential chemical mutagen for inducing variability in crop plants. The induction of biological damage in terms of instantaneous effects on seed germination, pollen fertility and seedling growth was envisaged by treating the seeds of two wheat (Triticum aestivum L.) varieties viz., HD-2285 and HUW-234 with 0.01%, 0.02%, 0.03% and 0.04% of SA. All the mutagenic treatments bring about dose dependent decline in seed germination, pollen fertility and seedling growth in M1 and M2 generations. The reduction was more prominent in M1 than M2 generation. For all the traits under study, var. HUW-234 was found to be more responsive than the var. HD-2285 to various doses of SA. The direction of shift in mean values for plant height indicated that negative micro- mutations outweighed the positive ones, while the reverse was true for number of tillers per plant, number of grains per spike, 100-grain weight and grain yield per plant. Coefficient of variation (CV) was recorded to be elevated in the mutagen treated population in both the generations for both the varieties, providing greater scope for attaining the favored selections. In M2 generation, the mean seed protein content of the mutants displayed no considerable variation from the controls. However, the coefficient of variation was of lesser magnitude, representing that the further improvement in this particular trait is pretty difficult to accomplish.
Wheat, sodium azide, biological damage, induced variability, yield components, protein content.
SA- sodium azide, RCBD- randomized complete block design, SE- standard error, CV- coefficient of variation, BOD- biochemical oxygen demand.
Wheat (Triticum aestivum L.) is the most important cereal crop both in regard to its antiquity and use as a source of human food. The significance of this crop is apparent from the fact that it is a staple food for more than one third of the world’s burgeoning population. Moreover, it is a key source of energy, proteins and nutritional fibers. As the global population is escalating progressively, the breeders are also trying to boost the productivity by making an arduous effort to bring out a balance between the population and productivity. The breeding of wheat by traditional methods has been practiced for centuries; however it has merely come to a stage where these methods are insufficient to make any further breakthrough to cope with the world’s mounting demand. The major constraint in the progression of improved varieties is the limited genetic variability among the existing wheat genotypes. However, Muller [28] opened a new era in the field of crop improvement and now mutation induction has become a reputable gizmo in the field of plant breeding to supplement the existing germplasm and improve cultivars in certain specific traits.
The prime strategy in mutation breeding has been to upgrade the well tailored plant varieties by altering one or two major traits which limit their productivity or augment their quality. Mutations have played a mammoth role in increasing the world food security by contributing significantly in the augmentation of crop production [18,37] . Mutation induction offers the prospect of inducing desired attributes that either cannot be found in nature or have been lost during evolution. Treatment with mutagens alters genes or rupture chromosomes. The mutation in genes crop up naturally as errors during DNA replication. Most of the errors are repaired; nevertheless some may pass to the subsequent cell division to become established in the plant off springs as spontaneous mutations. Gene mutations without phenotypic expressions are usually not recognized. Consequently, genetic variations appear rather limited and breeders have to resort to mutation induction [31] .
Chemical mutagenesis is regarded as an effective and central tool in improving the yield and quality characters of crop plants. The usefulness of mutation breeding in escalating the genetic variability for quantitative traits in various crop plants has been proved beyond doubt [7,27,40] . Sodium azide (NaN3) is one of the most potent mutagens exploited in crop plants. The mutagenicity is arbitrated through the formation of an organic metabolite which enters the nucleus, interacts with DNA and generates point mutations in the genome. To enhance the mutagenic efficiency of sodium azide particularly the metabolite, additional knowledge about the upshot of time, pH value, temperature, seed soaking and different concentrations is mandatorily required [16] .
Uniform and healthy seeds of wheat (Triticum aestivum L.) varieties HD–2285 and HUW-234 were presoaked in distilled water for 9 hours prior to mutagen treatment. Four different concentrations of sodium azide (0.01, 0.02, 0.03 and 0.04 %) were freshly prepared in phosphate buffer adjusted to pH-3. During the treatment, the flasks containing the solution and the seeds were recurrently shaken to ensure adequate aeration. Controls were maintained by presoaking the seeds in distilled water only. After completion of treatment period for 6 hours, seeds were scrupulously washed in running tap water to diminish the residual effect of the mutagen sticking to the seed coat. The experiment was layed out in randomized complete block design (RCBD) with split-plot arrangement. Three replications of 100 seeds each were sown for every treatment and controls for both the varieties to hoist M1 generation. The experiment was irrigated at suitable intervals in such a manner that the crop did not suffer from water stress. The M1 plants were harvested separately and the seeds were sown in next season in plant progeny rows to raise M2 generation. The distance between the seeds in a row and between the rows was kept as 10 and 25 cms respectively. Twenty five seeds from each treatment and controls were spread over moist cotton in petri-dishes and kept in B.O.D. incubator at 25±1oC temperature, in order to determine the percentage of seed germination and seedling growth. After ten days of sowing the seeds in petri-dishes, germination counts and growth observations were recorded on shoot and root length. Pollen fertility was determined by staining the pollen grains with 1% acetocarmine solution. For this purpose, 15 M1 and M2 plants at random were selected from each treatment and controls for both the varieties and finally 10 young flower buds from each plant were used for microscopic scrutiny. Pollen grains which absorbed stain and had a regular outline were considered as fertile, while shrunken, empty and unstained ones as sterile. The data collected for various quantitative traits viz., plant height, number of tillers per plant, number of grains per spike, 100-grain weight and grain yield per plant were analyzed statistically [35] to find out the mean, standard deviation (S.D.) and co-efficient of variation (CV). The protein content of the grains in M2 generation was determined by the method recommended by Lowry, et al. [25] .
The application of sodium azide on crops is effortless and inexpensive for the advancement of agronomic traits. It has long been used in various crops to improve their yield and quality traits, besides creating resistance against biotic and abiotic stresses. In the present experiment, both the varieties of wheat showed dose dependent reduction in various biological parameters such as seed germination, pollen fertility and seedling growth in M1 and M2 generations with increasing concentrations of SA. The inhibition in seed germination was recorded maximum 22.91% and 26.80% with 0.04% SA in the varieties HD-2285 and HUW-234 respectively in M1 generation [Table-1] , [Fig-1] . The seed germination percentage also decreased in M2, but it was markedly less as compared to M1 generation.
The diminution in seed germination has been elucidated due to interruption of physiological and biological processes indispensable for seed germination including enzymatic activity [22] . Usuf and Nair [38] reported that gamma rays hamper the synthesis and induce the degradation of existing enzymes involved in the formation of auxins which consequently reduce the germination of seeds. Azide’s mutagenic action might depend upon either metabolism or the state of DNA, i.e. S phase, in the embryonic shoot cells of the seed [23] . High reduction in germination percentage in SA treatments may be due to weakening and disturbances of growth processes. The greater sensitivity at higher doses of mutagens has been attributed to various factors such as changes in the metabolic activity of the cells [30] and disturbances of balance between promoters and inhibitors of growth regulators [26] .
In the control, the var. HD-2285 showed 98.36% of pollen fertility, while in the var. HUW-234, the fertility of pollen was recorded to be 98.48%. The maximum reduction in pollen fertility was found to be 15.51% and 17.95% with 0.04% SA in the varieties HD-2285 and HUW-234 respectively [Table-1] , [Fig-2] .
The reduction in pollen fertility was found to be more in M1 than in M2 generation, indicating the operation of recovery mechanism in the superseding period. The higher degree of pollen sterility was reported to be linked with meiotic abnormalities [15,29] because meiosis is more prone to conceivable type of turbulences [16] . Contrarily, Sato and Gaul [34] reported high pollen sterility coupled with low frequency of meiotic abnormalities in barley subsequent to EMS treatments. This was accredited to small untraceable deletions or gene mutations. The sterility induced by SA seems to be gene controlled in nature.
The percentage injury in seedling growth ranged from 7.24% to 23.98% in the var. HD-2285, whereas it ranged from 7.90% to 25.11% in the var. HUW-234 in M1 generation [Table-1] [Fig-3] The inhibition in seedling growth might have aroused due to the effect of mitotic impediment. The extent of decrease in seedling height was not uniform in the two varieties of wheat studied. It may be due to the uneven damage caused to the meristematic cells as a consequence of genetic injury. The badly damaged cells would produce only a few cell progeny and growth will recur from those cells which are least damaged genetically. Variation in auxin level [11] , change in the specific activity of quite a few enzymes [4] and physiological injury induced in the seeds and seedlings [12] were correlated with the lessening of seedling growth after mutagenic treatments. Evans and Sparrow [8] suggested that the chromosomal damage and inhibition of cell division are the chief causes of reduced seedling growth. Blixt [3] opined that the inhibition in seedling growth might be due to the gross injury caused at cellular level either due to gene controlled biochemical processes or acute chromosomal aberrations or both. Based on seed germination, pollen fertility and seedling growth, var. HUW-234 was found to be more sensitive than the var. HD-2285. The greater sensitivity at higher mutagenic level has been attributed to changes in the metabolic activity of the cells [24,37] . Varietal differences were also reported earlier with respect to mutagen sensitivity [2,13] . The sensitivity of an organism depends upon the type of mutagen employed and its genetic makeup [14] , amount of DNA and its replication time in the initial stages [39] , besides physiological factors such as pH, moisture, oxygen and temperature [19] . Comparative mutagenicity of SA in the two varieties viz., HD-2285 and HUW-234 reflects the difference in their genome architecture. Genetic difference even though very small can induce significant changes in the mutagen sensitivity, which in turn influences various plant characters [16] .
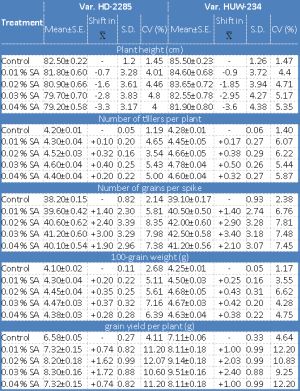
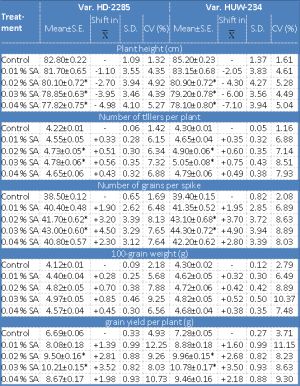
Data on the effect of SA on mean and coefficient of variation for different quantitative traits in M1 and M2 generations are presented in [Table-2] and [Table-3] . The mean values for plant height showed consistent dwindle with increasing concentrations of SA in both the generations for both the varieties. The maximum reduction in plant height was noticed in the var. HUW-234 in M2 generation [Table-3] . The extent of reduction in growth is related to the mechanism of action of a given mutagen. As a respiratory inhibitor, azide may hamper an energy system resulting in the inhibition of mitosis which can be associated with plant growth depression. These findings are in conformity with the preceding reported results [6,20,33] .
There was an increase in the mean number of tillers per plant and the number of grains per spike in all the mutagen treatments over the controls for both the varieties in M1 and M2 generations [Table-2] [Table-3] . For both these traits, 0.03% of SA treatment was more effectual in increasing the mean values from their respective controls. Number of tillers per plant and the number of grains per spike are the desirable attributes of a genotype from the breeding point of view in order to obtain the higher grain yield. Konzak, et al. [21] have reported parallel results by advocating that increase in the number of tillers is associated with an increase in the radiation intensity. However, quite contradictory results have been reported by Abrams [1] and Ghafoor and Siddiqui [10] , who observed an adverse effect of radiation on the number of tillers/plant. Such type of deviation might be due to different materials utilized and the agro-climatic conditions under which the experiments were conducted.
The weight of 100 grains is an extremely reliable measure of yielding aptitude in cereals. Even though the mean 100-seed weight displayed a positive shift, still the difference was not significant over the control in M1 generation for both the varieties. No significant improvement in grain weight was also reported in durum wheat after gamma rays treatment [32] . The grain yield of a crop is a composite trait, the result of interactions of various parameters. The data on grain yield per plant presented in [Table-2] and [Table-3] illustrated that there was a general increase in mean values for each treatment in M1 and M2 generations for both the varieties viz., HD-2285 and HUW-234. However, the mean in the treated population at the moderate (0.02, 0.03% SA) concentrations differed significantly from that of the control in M2 generation. The augment in the mean values of various traits may be due to the selection of normal looking plants in M1 which led to the abolition of aberrant plants and also due to the changes induced at genetic level. For all the quantitative traits studied in the present investigation, the coefficient of variability (CV) induced by the chemical mutagen was found to be larger in the mutagenized population for both the varieties. A non-linear relationship was observed between the mutagen concentrations and the variability induced for various traits. These results are in concurrence with the former report of Singh, et al. [36] who observed no linear relationship between the mutagen doses and the variability induced in urdbean. Induction of greater variability in polygenic traits might be due to the increased mutations and recombinations.
Grain protein content is considered to be a complex character governed by many genes located on several chromosomes [5,9] . The estimates for total grain protein content of the mutants isolated in M2 generation revealed that the mean protein content did not differ significantly as compared to controls [Table-4] [Fig-4] .
Furthermore, the coefficient of variation for seed protein content did not varied a great deal from the control plants, indicating that no further improvement in seed protein content of wheat is possible. Protein content is influenced by the interaction of genes and environmental factors. In the present study, variation in grain protein content in the mutant population may be due to change in environmental factors.
Genetic variability is indispensable for any crop improvement programme and its creation and management is vital to plant breeding. The nature and extent of genetic variability available within the species forms the basis of effectual selection for economic traits under improvement. The use of mutagens facilitate to comprehend the mechanism of mutation induction and to quantify the pattern of changes in different selected crop plants. In this study, the mean values shifted to positive direction for all the traits. The positive shift was more distinct at the moderate concentrations of the mutagen. The micro-mutations resulted in the release of substantial genetic variability in the mutagen treated population. Enlargement in range of variability for yield and yield contributing traits in M1 and M2 generations is indicative of the wider scope for selection. Estimates for seed protein content of the mutants in M2 generation demonstrated no significant fluctuation as compared to the controls. Moreover, the coefficient of variation for seed protein content has not greatly altered over the controls, indicating that additional improvement is rather intricate to achieve. Thus it is concluded that sodium azide at moderate (0.02% and 0.03%) concentrations appear to be the most effective treatment for inducing micro-mutations in yield component traits and succeeding selection in M2 population for these treatments would be exceptionally effective in bringing out the novel lines with high yield potential.
The authors are gratified to the Chairman, Department of Botany, Aligarh Muslim University, Aligarh, India and Principal, Govt. Degree College (Boys), Anantnag, Jammu and Kashmir, India for providing all the requisite infrastructural support with regard to these experiments.
[1] Abrams R. (1957) J. Agr. Univ. Puerto. Rico., 41, 242-258.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[2] Akbar M., Inoue M. and Hasegawa H. (1976) The Nucleus, 13, 25-29.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[3] Blixt S. (1978) Agr. Hort. Genet., 25, 121-130.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[4] Cherry J.H., Hagemann R.H. and Hanson J.B. (1962) Rad. Res., 17, 740-754.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[5] Coffman W.R. and Juliano B.O. (1979) IAEA, Vienna, 261-276.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[6] Conger B.V. (1973) Rad. Bot., 13, 375-379.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[7] Das S.V. and Chakraborty S. (1998) Plant Sci., 4, 277-281.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[8] Evans H. J. and Sparrow A. H. (1961) Brookhaven Symposia in Biology, 14, 101-122.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[9] Frey K.J. (1977) Protein of oats. Z. Pflanzenzucht., 78, 185-215.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[10] Ghafoor A. and Siddiqui K.A. (1976) Pak. J. Botany, 8, 1-7.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[11] Goud J.V. and Nayar K.M.D. (1968) Mysore J. Agric. Sci., 11, 53-55.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[12] Ignacimuthu S. and Babu C.R. (1988) Indian J. Genet., 48(3), 331-342.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[13] Irfaq M. and Nawab K. (2003) Asian J. Plant Sci., 2(13), 999-1003.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[14] Kaul M.L.H. (1988) Male Sterility in Higher Plants. Springer Verlag, Germany.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[15] Khan S. and Wani M.R. (2005) Adv. Plant Sci., 18(2), 533-535.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[16] Khan S. and Goyal S. (2009) African J. Plant Sci., 3(8), 174-180.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[17] Khan S., Al-Qurainy F. and Anwar F. (2009) Environ. Int. J. Sci. Tech., 4, 1-21.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[18] Kharkwal M.C. and Shu Q.Y. (2009) The Role of Induced Mutations in World Food Security. In: Shu Q.Y. (ed.), Induced Plant Mutations in Genomics Era. Food and Agricultural Organization of the United Nations, Rome, Italy, 33-38.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[19] Konzak C.F., Nilan R.A., Wagner J. and Foster R.J. (1965) Rad. Bot., 5, 49-70.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[20] Konzak C.F., Niknejad., Wickham M.I. and Donaldson E. (1975) Mut. Res., 30(1), 55-62.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[21] Konzak C.F., Woo S.C. and Dickey J. (1969) Wheat Information Service, 28, 10.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[22] Kurobane I.H., Yamaguchi H., Sander C. and Nilan R.A. (1979) Env. Exp. Bot., 19, 75-84.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[23] Kleinhofs A., Owais W.M. and Nilan R.A. (1978) Mut. Res., 55, 165-195.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[24] Krishna G., Shivshankar G. and Nath J. (1984) Environ. Exp. Bot., 24, 197-205.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[25] Lowry O.H., Rosebrough N.J., Farr A.L. and Randall R.J. (1951) J. Biol. Chem., 193-265.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[26] Meherchandani M. (1975) Rad. Bot., 15, 439-445.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[27] Mostafa G. G. (2011) Int. J. Plant Breed. Genet., 5, 76-85.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[28] Muller H.J. (1927) Science, 66, 84.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[29] Muthusamy A. and Jayabalan N. (2002) Indian J. Genet., 62(2), 187.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[30] Natarajan A.T. and Shiva Shankar G. (1965) Z. Vererburgstehre, 43, 69-76.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[31] Novak F.J. and Brunner H. (1992) Int. Atomic Energy Agency Bulletin, 4, 25-32.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[32] Potdukhe N.R., Wanjari S.S. and Raut S.K. (1994) Agricultural Science Digest, 14, 121-125.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[33] Sander C., Kleinhofs A., Konzak C.F. and Nilan R.A. (1972) Barley Genet. Newslett., 2, 71-72.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[34] Sato M. and Gaul H. (1967) Rad. Bot., 7, 7-15.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[35] Singh R.K. and Chaudhary B.D. (1985) Biometrical Methods in Quantitative Genetic Analysis. Kalyani Publishers, Ludhiana, India.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[36] Singh V.P., Singh M. and Lal J.P. (2000) Indian J. Genet., 60(1), 89-96.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[37] Srivastava P., Marker S., Pandey P. and Tiwari D. K. (2011) Asian J. Plant Sci., 10(3), 190-201.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[38] Usuf K.K. and Nair P.M. (1974) Rad. Bot., 14, 251-256.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[39] Varughese G. and Swaminathan M.S. (1968) Indian J. Genet., 28, 150-165.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[40] Wani M.R., Khan S. and Kozgar M.I. (2012) Crop Research, 43(1-3), 189-193.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
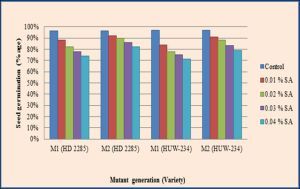 | Fig. 1- Effect of SA on seed germination in M1 and M2 generations of wheat (Triticum aestivum L.). |
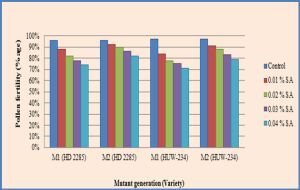 | Fig. 2- Effect of SA on pollen fertility in M1 and M2 generations of wheat (Triticum aestivum L.). |
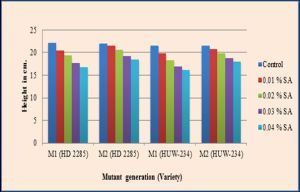 | Fig. 3- Effect of SA on seedling growth in M1 and M2 generations of wheat (Triticum aestivum L.). |
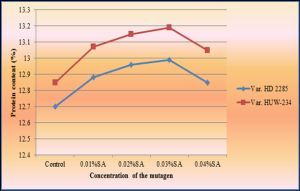 | Fig. 4- Estimation of seed protein content of mutants isolated in M2 generation of wheat (Triticum aestivum L.). |
 | Table 1- Effect of sodium azide (SA) on seed germination, pollen fertility and seedling growth in M1 and M2 generations of wheat (Triticum aestivum L.) |
 | Table 2- Estimates of mean values, shift in ( |
 | Table 3- Estimates of mean values, shift in ( |
 | Table 4- Estimates of mean, Shift in ( |