ISSN : 0976-9927
EISSN : 0976-9935

ODO G.E.1*, DIBUA U.E.M.2
1Department of Zoology and Environmental Biology, University of Nigeria, Nsukka, Enugu State, Nigeria.
2Department of Microbiology, Faculty of Biological Sciences, University of Nigeria, Nsukka, Enugu State, Nigeria.
* Corresponding Author : odogreg@yahoo.com
Received : 07-08-2013 Accepted : 09-09-2013 Published : 10-10-2013
Volume : 4 Issue : 2 Pages : 95 - 102
J Fish Aquaculture 4.2 (2013):95-102
DOI : http://dx.doi.org/10.9735/0976-9927.4.2.95-102
Aspects of food and feeding profile and haematoplasma chemistry of the Protopterus annectens were studied over 12 months. The 1131 fish (30.6-46.1cm TL) examined for food composition 22 (2.04%) had full stomachs, 513 (49.81%) empty and 496 (48.15%), partially-filled stomachs. It fed on fish (98.60% IFD) while frog eggs, gastropods, cephalopods, tadpole, insect remnants and seeds were of minor importance, each forming <1.00 % IFD of diet. The mean blood parameters obtained were Haemoglobin (Hb) 33 ±0.61g/dl; Packed Cell Volume (PCV) 26.00 ± 2.73%; Red Blood Cells (RBC) 4.29 ± 0.32 x 106 cells/mm3; White Blood Cells 2.40 ± 0.34 x 104 cells mm3; Erythrocyte Sedimentation Rate (ESR) 29.33 ± 2.72µm; Mean Cellular Haemoglobin Concentration (MCHC) 32.00 ± 2.39%; Mean Cellular Haemoglobin (MCH) 19.00 ± 2.55pg; Mean Cellular Volume (MCV) 60.67 ± 2.57µm3;.Biochemical profile gave: plasma sodium 11.66 +/- 1.55 Mm, plasma potassium 16.64 +/- 3.27 mM, plasma chloride 2.76 +/- 1.16 mM, plasma magnesium 4.38 +/- 1.65 mM, plasma phosphorous 362.20 +/- 183.73 mM, plasma glucose 39.69 +/- 1.65 mg/dl and plasma albumin 4.2 +/- 0.92 mg/dl. Blood Group 0- (83%) and 0+ (17%), Genotype AS (86%) and AA (14%).Digestive enzymes assays revealed an array of glycosidase; proteases and lipases.
Protopterus annectens, Feeding habits, digestive enzymes, haematological profile, Anambra River, plasma chemistry, Nigeria.
The Nigerian flood river system supports considerable stocks of the African lung fish, P. annectens which constitute significantly proportion of the canoe landings of artisanal fishermen. The P. annectens is widely distributed in the swampy and brackish waters or rivers during wet seasons. The fish, P. annectens is economically valuable and is the only species of the primitive family, lepidosirenidae found in West Africa fresh waters [1] . The fish is characterized by a pair of lungs and strong solid teeth with powerful bony-ridge [2] . The feeding adaptations (dentitions, gill rakers and gut systems of the fish have been described [3,4] . Studies on the biology of P. annectens include those of Reed, et al [1] and Adeyemi, et al [5] among others. Adeyemi [6] reported that P. annectens of Kogi State Bassa Lake, Gbedikere fed almost exclusively on fish parts, sand, insect parts The study of dietary habits of fish based on stomach content analyses is widely used in fish ecology as an important means of investigating relationships in aquatic communities and formulating management strategy options in multispecies fisheries. Notwithstanding the P. annectens enormous importance, knowledge on its food and feeding profile in Anambra River is scarcely known. Svobodova, et al [7] reported that Ichthohaematology is very useful in the determination of disease condition of fish, toxic effects of substances, evaluation of fish conditions as well as suitability of feeds and feed mixture. Thus the knowledge of haematological profile of normal fish species is very important since variations in the values of blood parameters indicate the health status of the fish. A fish species haematology also reflects its dietary sufficiency and degree of protein content.
The haematological profile of some tropical African fish species have been reported, namely, C. isheriensis [8] , C. gariepinus, H. longifihs and C. nigrodigitatus [9,10] , Heterobranchus bidosarhis [11] , O. niloticus [12] , H. fuciatus and T. zilli [13] , S. melanotheron [14] , P. obscura [15] . There are no reports on the haematological profile of P. annectens from Anambra River, Nigeria. Hence the need to study the heamatological profile to provide some useful information on this aspect of its biology. Thus, this study, which forms part of a larger and on-going observation on fish species load of the river, is an attempt to fill this information gap and addresses aspects of the qualitative and quantitative composition of food items, ontogenetic changes in food and feeding habits, Sex-dependent changes in the food composition, sex-dependent variation in feeding intensity, monthly changes in stomach repletion and monthly dynamics in food richness. Also aspects of the haematological, plasma chemistry, serological profiles, the occurrence, distribution and relative activities of glycosidase, proteases and lipases in the different gut regions of P. annectens will be addressed.
The study area was the Anambra River about 14014km2 [16] . The Anambra River [Fig-1] is about 207.4 km in length; it rises from the Ankpa hills (ca. 305-610m above sea level), flows in southerly direction through a narrow trough that gradually broadens as it courses down. It crosses the Kogi/Anambra state boundary a bit north of Ogurugu, then meanders through Ogurugu and eastern part of Otuocha from there it flows down to join River Niger at Nsugbe near Onitsha. The basin lies between latitude 6° 861 N and 7° 31 N longitude 6° 351 and 7° 401 east of the River Nigeria. There are two main seasons, the dry period of December-March and the rainy season (April-September/October) approximately corresponding to the dry and flood phase, respectively of the hydrological regime [16] . The vegetation is somewhat transitional between the Equatorial Rain Forest and the Savannah grass types. Also the ecology and productivity of the river basin have been extensively studied [17] .
One thousand one hundred and thirty one (1131) mature, averaged sized, live fishes of P. annectens were obtained from Anambra River through local fishermen. The fishes were transported to the Laboratory and kept in an aquarium supplied with filtered and aerated tap water for two weeks for acclimatization to laboratory conditions. During this period the fishes were fed to satisfaction twice daily with 400g of crude protein pellets. All the fishes were considered healthy on the basis of their appearance and absence of obvious sign of disease condition. No sex selection was made. The standard length of each fish was taken using a measuring ruler. The standard length is the distance between the mouth tip and the beginning of the caudal fin. Each fish was also weighed using a weighing balance and the values recorded in grams They were preserved for 2-5 days in deep freezer (at -10°C) pending further examination. Specimens were later dissected and the stomachs removed and slit open. The degree of fullness of each was estimated by an arbitrary 0-20 points scale allotted to empty, ¼ full, ½ full, ¾ full and fully distended stomachs respectively. The percentage of empty stomachs (ES), full stomachs (FS), partially filled stomachs (PS) (i.e. ¼ - ¾ full) and average stomach fullness (AS) (mean points per stomach) were used to evaluate patterns in feeding activity. The stomach contents of each specimen were placed in a Petri dish and aggregates dispersed with a small amount of water prior to macroscopic and microscopic examinations. The contents were sorted, identified and the importance of each was assessed by the relative frequency (RF) and percentage points (PP) by King [18] in trophic studies on Liza species.
The integrated importance of each item was then expressed by an index of food dominance (IFD) according to the formula:
(1)
This index ranges from 0 to 100%. Items with IFD ≥ 10% were arbitrarily considered as dominant food items; those with IFD 1-9.9% as secondary and those with IFD < 1% as incidental. The use of IFD to establish overall food preponderance is adequate as it incorporates the RF and PP data, thus minimizing the bias characteristics of cases in which results from different analytical methods are independently interpreted. Food richness was defined as the number of major items in the diet, the IFD data were used to compute diet breath based on Shannon’s function, H’ [18,19] . Feeding intensity and food composition data were analyzed by d-statistic and T-test [20] . Sixty live specimens (TL 32-41 cm) were kept undisturbed in large glass aquaria (120 liter capacity) supplied with filtered and aerated tap water for 2 weeks of acclimation prior to haematological studies. Blood was taken each fish using separate heparinized disposable syringes and hypodermic needles. Haematocrit (PCV) was measured after centrifugation at 15000 rpm (MSE Micro centrifuge). The method of blood analysis described by Svobodova, et al [7] was followed. Blood cell count (erythrocytes and leucocytes) was carried out in an improved Neubaeur haemocytometer using a modified Yokoyama diluting fluid. The basic erythrocyte indices, Mean Cell hemoglobin concentration (MCHC), Mean Corpuscular Volume (MCV) and Mean Corpusclar Haemoglobin (MCH) were computed from haemoglobin values and erythrocyte count. Determination of the biochemical components (Na, Cl, Ca, Mg, K, P) ion concentrations were carried out using the methods of Svobodova, et al [7] . The plasma electrolyte (plasma sodium and potassium) were determined by flame photometry using acorning 400 photometer. Plasma magnesium was determined by using MODEL 200A flame chloride done by titrimetric method described by Schales and Schale [21] ; while plasma phosphorus was determined by spectrophotometer method [22] . Blood grouping was performed with by the test tube techniques while the genotype was determined by electrophoresis steps [23] . During this period, the fish were fed to satiation twice daily with 380g/kg of crude protein pellets. All fish were considered healthy on the basis of their appearance and absence of obvious signs of disease. No sex selection was made. The fish were caught individually using hand net and stunned by a blow on the head. Blood was collected from the caudal vein of each fish using separate heparinized disposable syringes and hypodermic needles. Sixty adult P. annectens specimens (TL 42.1-49.6cm) were kept unfed for 72 hrs. inside out door concrete cisterns in order to bring them to a similar physiological condition as well as to ensure the emptiness of the entire gut. They were anaesthetized with benocaine and dissected to remove the entire gut, later separated into the anatomically distinct regions The different gut regions were pooled and homogenized; the homogenates were then centrifuged at 1200 rpm at 4°C. The supernatants were used as crude extracts. The methods of Olatunde, et al [24] , was used for glycosidase determination while quantitative assays were conducted using the dinitrosalicyclate (DNS) methods [25] . Qualitative and quantitative assays of proteases followed the method of Baloggun and Fisher [26] . The methods described by Ogunbiyi and Okon [27] were used to determine lipase activityThe coefficient of regression analysis was carried out between the various haematological parameters and standard length. These inferences were then analyzed for statistical significance by student’s t-test (p=0.05) Food richness was defined as the number of major items in the diet, IFD data were used to compute diet breath based on Shannon’s function, H’ [18,19] Feeding intensity and food composition data were analyzed by d-statistic and T-test [20] . The number of specimens examined per month and their size ranges are presented in [Table-1] . Data for corresponding months were pooled together for sorting and determination. Food composition and sex ratio were analyzed by students-test and X2 test, respectively [20] . Differences were considered significant at 5%.
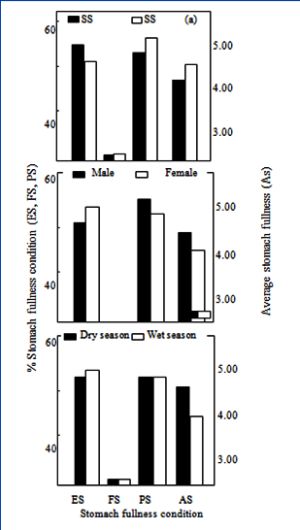
The mean values of water analysis during the present study are presented in [Table-2] . Also of the 1131 specimens of P. annectens (30.6-46.1cm TL) examined for food composition [Table-1] 22 (2.04%) had full stomachs 513 (49.81%) empty and 496 (48.15%), partially-filled stomachs. In order to assess the ontogenetic changes in food and feeding habits of P. annectens, specimens were categorized into two size-groups (small-sized group (SSG<40cm TL) and large-sized group (LSG≥40cm TL)). These two size-groups were chosen since according to King (unpubl.), P. annectens of the SSG is sexually immature while those of the LSG comprise mature fishes [Fig-1] Illustrates the indices of stomach fullness of the SSG (size range 30.6-39.9cm TL; n = 535) and LSG (size range 40-46.1cm TL; n = 487) examined. There was no significant difference in FS and PS between the size groups (d-test: P>0.05 in each case) however, ES was significantly higher in the SSG (d = 1.988, p<0.05) while As was higher in the LSG (t = 1.775, P<0.05). [Fig-2] depict variations in indices of feeding activity ES -% empty stomachs; FS = % full stomachs; PS-% partially filled stomachs; AS-average stomach fullness) of P. annectens in relation to size (a) (SGG-small-sized group; LSG-large-sized group), sex (b) and season (c). The overall stomach content of P. annectens [Table-3] revealed that14 major items were ingested, of these 2 were of primary importance and 12 of incidental importance.
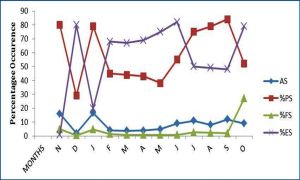
It fed predominantly on fish (98.60% IFD) while frog eggs, gastropods, cephalopods, tadpole, insect remnants and seeds were of minor importance, each forming <1.00 % IFD of the diet. Fish consumed were dominated by fry and eggs and closely followed by the composite of frog, fish parts and tadpole and this accounted for only 0.68% IFD of the diet. Cephalopods in the diet were represented by sepia. [Table-4] illustrates the ontogenetic changes in the trophic spectra of the two size-groups of P. annectens. There was no size-dependent variations in food richness although slight differences were observed in the quantitative food composition of the size-groups. For instance, mysids and insect remnants were not ingested by the SSG while amphipods were absent from the di et of the LSG. Similar trends occurred in the rank-order of the IDF of the food items (Spearman rank correlation: rs-0.823, p<0.02) of the two size-groups although there were marked differences in the proportion of some of the items. There was an increase with fish size of the IFD of shrimps and cephalopods and a decrease in that of mysids and fish scales. No marked size-based changes were apparent in the relative importance of amphipods, unidentified fish, fish fry, fish eggs, tadpoles and frog eggs. Indices of diet breadth were higher in the SSG than in the LSG [Table-4] , indicating an increasing food specialization with fish growth Sex-dependent changes in the food composition of P. annectens are summarized in [Table-5] . Food richness was higher in females than males by a factor of 6. Fish, cephalopods, unidentified gastropods, insect remnants and seeds were not ingested by males while the complete array of items shown in [Table-4] was consumed by the females. Although both sexes exhibit similar patterns in the rank-order of the IFD of the food items (rs=0.822, p<0.002), the proportions of some of them were different. A total 1131 specimens, comprising 314 males (size range 30.6-46.2 cm TL) and 714 females (size range 30.6-46 cm TL) were examined for sex-dependent variation in feeding intensity. The stomach repletion of both sexes [Fig-3] showed no significance in FS (d-test: p<0.05) between them. However, ES was significantly higher (d = 2.412, p<0.02) in females than males while males had significantly higher PS (d = 2.188, p< 0.05) and AS (t = 33.000, p<0.01) than females. The monthly changes in stomach repletion [Fig-3] showed that peak PS, ES and AS occurred in November and January, these coinciding with the months with lowest ES. These results indicate high feeding intensity in November, January and July-October while low feeding intensity occurred in December and February-May. A total of 396 specimens (37.0-46cm TL) were examined in the dry season and 632 (30.6-46 cm TL) in the wet season. The seasonal variation in stomach fullness conditions is presented in [Fig-2] .
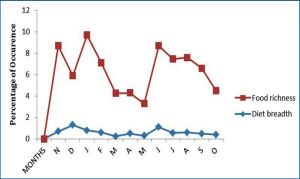
There was no significant seasonality in ES and PS of the fish (d-test: p>0.05 in each case). However, there was a significant dry season increase in FS (d = 2.778, p<0.001) and AS (t = 10.733, p<0.001), thus suggesting that feeding intensity was higher in the dry seasons than during the rains. Males had significantly higher IFD of mysids and nematodes than females, while females had higher IFD of shrimps than males. There was no variation with sex of the IFD of amphipods unidentified fish, fish scales and fish eggs. Diet breadth was not markedly different in both sexes [Table-4] . Monthly dynamics in food richness [Fig-4] ranged from 3 in May to 9 in January and August. High values (7-9) were recorded in November, January, February and June-September while low values (3-5) was obtained for all other months. Fish predominated in all months with peaks in November, January, March-April and August-October. Crustaceans were of primary importance only in December and June; it was of secondary importance in January-May and July while in November and August-October, it was of incidental importance. The insect and seed components of the diet were consumed as incidental items in January and August respectively. Mollusks occurred as incidental food items in November, January and June and insect remains in August and September. The monthly rhythms in diet breadth [Fig-4] showed high variability with values ranging from 0.32 in March to 1.31 in June; it was less than 1 in all months except December and June when values exceeded 1, [Table-6] .
The composite diet data for the two main seasons [Table-7] showed that the wet season food richness was slightly higher than the dry season’s value. The quantitative food compositions portrayed high similarity in both seasons apart from the exclusion of unidentified gastropods and insect during the rains and fish larvae and cephalopods during the dry season. Similar seasonal trends occurred in the rank-order of the IFD of the food items (rs = 0.808, p<0.002), although there were differences in some of their relative proportions. There was a marked wet season increase in the IFD of mysids and a dry season increase in that of shrimps, unidentified fish and fish eggs. No significant seasonality occurred in the IFD of amphipods, fish scales, fish intestinal parts and cephalopods. Diet breadth was slightly higher in the dry season than during the rains. The mean values for the blood parameters are presented in [Table-8] .
There were intra-species variations in the blood values as indicated by the wide ranges of some parameters in the study. The highest range in the parameters was recorded in PCV while the lowest range was observed in RBC [Table-9] . The levels of correlation among the the different blood parameters are presented in [Table-10] . There is very high and positive correlation between RBC and Hb, PCV and Hb, ESR and MCH, MCV and MCH, MCV and MCHC and between MCV and ESR at 0.01% level of significance. But a low positive correlation occurs between Hb and MCH, WBC and ESR at 0.05% level of significance. A negative correlation exists between RBC and ESR, RBC and MCH and between RBC and MCV. The same negative correlation also occurs between Hb and ESR. The results of Blood Group and Genotype obtained were recorded in [Table-11] . Majority of the fish falls within the blood Group O with the prevalence of 83% and the prevailing Genotype being AS with the prevalence of 96%. [Table-12] shows the various enzymes detected in the different regions of the P. annectens gut, their distribution and activity varying along the gut length. A variety of glycosidase was detected [Table-12] . Cellulase activity was recorded only in the pyloric caeca [Table-12] . The protein hydrolyzing enzymes found in the stomach are pepsin-like while those in the pyloric caeca are alkaline proteases, possibly trypsin and /or chymotrypsin.
The proportion of empty stomachs of P. annectens observed here is high. The large proportion of empty stomachs in specimens from this flood plain system could be due to the regurgitation of food when the fishes were caught in gill-nets although this is unlikely for all specimens. Another possibility is that feeding is restricted to certain periods of the day and the specimens examined were probably caught during low feeding activity. It is also attributable to a generally low feeding activity of the population which may be induced by low availability and abundance of specific food resources in the estuary. The latter assertion, however, requires further investigation. This study revealed that the feeding intensity of P. annectens increased with fish size. This is not consistent with the idea of a negative correlation between feeding intensity and fish size relative to metabolic rates [2] . The feeding intensity of male P. annectens was significantly higher than of the female; the precise reason for this difference is uncertain but could be linked to sex-related differential energy requirements. The seasonality in stomach fullness conditions of P. annectens indicated a higher feeding intensity in the dry season than the rains; a pattern analogous to that of mormyrid, Brienomyrus branchyistius (Gill) in Nigeria [19] .
According to Baloggun and Fisher [26] the peak abundance of shrimps in Nigerian coastal waters is during the dry season (November-February). Therefore, the dry season increase in feeding intensity of P. annectens in Anambra flood plain system could be linked to the increase abundance of one of its principal food items-the fish species. This study showed that the relative importance of large-sized items such as shrimps and cephalopods increased with fish size while that of small-sized items such as mysids and fish scales decreased. The inherent increase in mouth gape with body growth of the fish probably permits this prey-size related feeding pattern by P. annectens. Ontogenetic variation in relative importance of fish diet (as observed in P. annectens) may result from changes in food predilection and/or foraging ability for the preferred food items. The ecological significance of the diversification in fish with growth is that it minimizes intraspecific competition and offers a wider spectrum of food resources for exploitation by the species [19] ). The food richness and diet breadth of P. annectens from Anambra flood plain system did not change with growth. These contradict [28] the assertion that as most fish’s food spectrum widens, food richness increases with growth. The observed sex.-related divergence in food habit of P. annectens probably reduces intersexual competition for food. Generally, the ranges of blood parameters determined for P. annectens are similar to those reported for Africa fresh water catfishes and tilapias except for those of erythrocyte count (RBC), haemoglobin concentration Hb and leucocytes count (WBC), which are wider in P. annectens. The mean haematocrit value (PCV) of P. annectens is comparable to those of Heterotis niloticus [12] and many Africa catfishes like C. isheriensis [8] C. gariepinus [9] and Chrysichthyes [9] . The mean PCV and WBC values of P. annectens are higher than the values reported for these mentioned catfishes [9] . Similarly the Haemoglobin (Hb) value, Red Blood Cell count (RBC) value and PCV of P. annectens are much higher than those reported for Sarotherodon melanotheron [14] but the mean cellular Heamoglobin (MCH), mean cellular volume (MCV) and white blood cells (WBC) of P. annectenss are lower than those reported for S. melanotheron, while the mean cellular haemoglobin concentration (MCHC), values of both fishes are the same. Furthermore the RBC count, Hb and MCHC values of P. annectens are much higher than those recorded for moron fish. The mean WBC of both fishes are in the same range the MCV, MCH and PCV values of the moron fish are higher than those of P. annectens [29] . However, the haematological features of P. annectens can also be compared to those of other vertebrates especially mammals. The RBC count of P. annectens is much lower than those recorded for monkeys, pigs, sheep and horse whose values for normal individuals ranges from 8.0 to 90 x 106/mm3, from 5 to 10 x 106/mm3, from 8 to 14 x 106/mm3 and from 5 to 11.0 x 106/mm3, respectively but it is in the same range with that of man, goat and cattle which has their values ranging from 4.2 to 6.2 x 106/mm3, from 4.0 to 12.0 ´106/mm3 and 4 to 11.0 x 106/mm3, respectively [29] . In the same way its Hb values falls in the same ranges with those of these mammals with the exception of man whose normal range is much higher than all, ranging from 12 to 17g/dl for a normal individual. The MCV of this fish is much higher than those of some of these mammals such as goat whose range is from 15 to 22µm3, sheep from 28 to 34µm3, cattle from 44 to 59µm3, horse from 26 to 58µm3. But the fish MCV is in the same range with those of monkey from 50 to 105µm3, pig from 50 to 62µm3 and man from 62 to 92µm3. The MCH of P annectens is the same range with that of all the above mammals with the exception of man. Then it is interesting to note that the MCHC of this fish has the same range with those of all the above mentioned mammals including man. The PCV of the fish is lower than those of monkey, cattle, sheep and man but has the same range with goat and horse. The differences in the range values of these various species of animals are ascribed to the differences in their genetic composition and varied environmental influence on each species. The high values of the erythrocyte count and haemoglobin concentration reflect a high oxygen carrying capacity of the fish blood which is in correlation with the haemoglobin concentration and fish activity. Lenfant and Johansen [30] reported that haemoglobin concentration is higher in fishes capable of aerial respiration more especially in lung fishes. Hence the high Hb values of protopterus are indicative of its air breathing character and high activity. From MCHC results, it is evident the fishes were healthy in terms of iron sufficiency and plasma protein, glucose and albumin values of 6.41±1.69g/l, 13.48, 4.16±0.92 were obtained for Labeo coubie. These were relatively low when compared with the value Ayotunde [31] reported for Heterotis niloticus: plasma protein (57.10±4.7g/l) and plasma glucose (61.46±5.29mg/l) while a higher value (7.53±1.83 mg/l) of plasma albumin was obtained for M. electricus by Ayotunde and Ochang [32] than that obtained in this study (4.16±0.92). Ayotunde [31] reported an almost similar value for H. niloticus (4.76±0.83 mg/l). The result for Blood group and Genotype showed a dissimilar antigen reaction to those shown in human blood. The predominant blood group for Protopterus is 0- (83%) while the predominant genotype is AS (86%). The results of the experimentation portrays that the haematological characteristics of the fish resemble that of human though with some slight differences, but it could be used in determining the health status of the fish for its management under captive rearing. The knowledge of the haematological profile of P. annectens indicates the strong viability of the fish, its dietary sufficiency and stable physiological response to environmental stress. The relatively high activity levels of protein diet in fish body parts are not surprising in view of the large proportion of zooplankton in the diet. Pepsin would hardly be expected to occur in the two distal gut regions since it is active only in strongly acid media found in the stomach. Lipase distribution and activity along the entire gut was also reported in Clarias isheriensis by Fagbenro [12] . This worker showed that P. annects is well equipped to digest its food components available in its diet.
[1] Reed W., Burch J., Hopson A.J., Jennes J. and Yaro I. (1967) Fish and Fisheries at Northern Nigeria, 226.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[2] Lagler K.F., Bardach E.J., Miller R., May R. and Passino D.R.(1977) Ichthyology, John Wiley and Sons, New York, 506.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[3] Ipinjolu J.K., Malami G.Z., Hassan W.A. and Magawata I. (2004) Annual Conference of Zoological Society of Nigeria, Institute of Development Research, Ahmadu Bello University, Zaria.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[4] Malami G.Z., Ipinjolu J.K., Hassan W.A. and Yaro I. (2004) Annual Conference of Zoological Society of Nigeria, Institute of Development Research, Ahmadu Bello University, Zaria.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[5] Adeyemi S.O., Bankole N.O. and Adikwu A.I. (2009) Continental Journal of Biological Sciences, 2, 7-11.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[6] Adeyemi S.O. (2010) Continental Journal of Biological Sciences, 6(2), 69-74.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[7] Svobodova Z., Prouvda D., Palackova J. (1991) Unified Methods of Haematological Examination of Fish, Research Institute of Fish Culture and Hydrology, Vodany, Czechoslovakia, 31.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[8] Kori- Siakpere O. (1985) Journal of Fish Biology, 27, 259-263.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[9] Erondu E.S. and Nwadukwe N. (1993) Journal of Applied Ichthyology, 9, 250-256.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[10] Sowunmi A.A. (2003) The Zoologist, 2, 96-122.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[11] Fagbenro O.A., Adedike C.O., Owoseeni E.A. and Yaoundé E.O. (1993) Tropical Zoology, 6, 67-79.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[12] Fagbenro O., Adedire C.O., Ayotunde E.O. and Famino E.O. (1999) Tropical Zoology, 13, 1-9.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[13] Egwunyenga A.O., Nmorsi O., Igbinosun A.F. (1991) Biology Science Research, 11, 361-365.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[14] Gabriel U.U., Anyanwu P.E. and Akinrotimi A.O. (2007) Journal of Fish Biology, 2(2), 186-189.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[15] Odo G.E., Onoja S.U., Onyishi G.C. (2012) International Journal of Fisheries and Aquaculture, 8, 154-169.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[16] Azugo W.I. (1978) Ecological Studies of the Helminth Parasites of the Fish of Anambra River System, M. Phil Thesis, University of Nigeria, Nsukka, Nigeria, 178.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[17] Awachie J.B.E. and Ezenwaji H.M.G. (1981) CIFA Technical Paper, 8, 212-233.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[18] King R.P. (1988) Revision Hydrobiology Tropics, 2(1), 63-70.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[19] King R.P. (1989) Cybium, 13(1), 25-36.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[20] Bailey N.T.J. (1994) Statistical Methods in Biology, 3rd ed., Cambridge University Press, 255.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[21] Schales O. and Schale S.S. (1941) Journal of Biological Chemistry, 140, 879-888.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[22] Fiske C.H. and Subbarow Y. (1956) Journal of Biological Chemistry, 66, 357-403.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[23] Delaney J.W. and Garratty G. (1969) Handbook of Haematological and Blood Transfusion Technology, Butterworths and Co. Ltd., London, 165.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[24] Olatunde A.A., Ukoha A.I., Oguntayo B.F. (1988) Archive for Hydrobiologie, 112(1), 107-113.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[25] Plummer D.I. (1978) An Introduction to Practical Biochemistry, 2nd ed., McGraw Hill Book Company, London, 283.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[26] Baloggun R.A. and Fisher O. (1970) Comparative Biochemistry and Physiology, 33, 813-820.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[27] Ogunbiyi A.O. and Okon E.E. (1976) Comparative Biochemistry and Physiology, 55, 359-361.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[28] Nkol’ski GV (1963) Zoologica, 46, 145-153.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[29] Mitruka B.M., Rawsley H.M., Vadehra B.V. (1977) Clinical Biochemical and Haematological reference Values in Normal Experimental Animals, Masson Publishing USA, Inc., New York. 271.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[30] Lenfant C. and Johansen K. (1972) Respiratory Physiology, 14, 211-218.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[31] Ayotunde E.O. (1998) Haematological and Serological Characteristics of Heteortis niloticus (C & V) 129,0 Osteogglossidae, M. Agric. Tech. Thesis, Federal University of Technology, Akure, 82.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[32] Ayotunde E.O. and Ochang S.N. (2004) Global Journal of Agricultural Sciences, 3(1 & 2), 5-14.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[33] Balogun J.K. (1980) A Biological Survey of Fishes of Epe Lagoon, Nigeria, M.Sc. Dissertation, University of Lagos, Nigeria. 164.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
 | Fig. 1- Map showing the location of study area |
 | Fig. 2- Variation in indices of feeding activity |
 | Fig. 3- Monthly variations in the stomach fullness condition of P. annectens in Anambra River |
 | Fig. 4- Monthly variation in food richness and diet breadth of P. annectens in Anambra River |
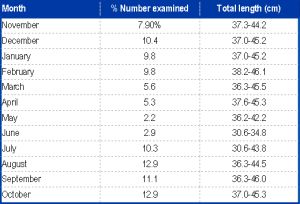 | Table 1- Percentage monthly numbers and size of P.annectens examined for food (November 2010 - October 2011) |
 | Table 2- Water quality parameters |
 | Table 3- overall trophic spectrum of P. annectens |
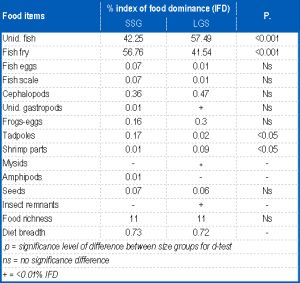 | Table 4- The trophic spectra of the small-sized group (SSG) and large-sized group (LSG) of P. annectens |
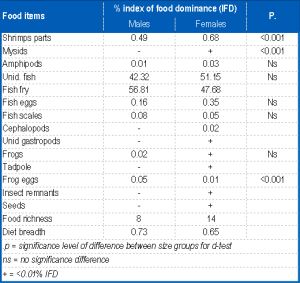 | Table 5- Sex dependent variation in the food composition of P. annectens |
 | Table 6- Monthly variations in the % index of food dominance (IFD) of P. annectens |
 | Table 7- Seasonal variation in the food composition of P. annectens |
 | Table 8- Summary of haematological parameters and plasma chemistry of P. annectens |
 | Table 9- Linear Regression Analyses of the Blood Parameters with Standard Length and Weight |
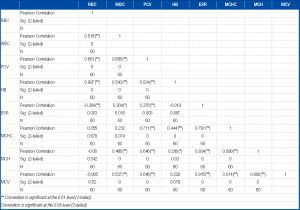 | Table 10- Correlation analysis of blood parameters of P. annectens |
 | Table 11- Blood Group and Genotype for the fishes (n=60) of P. annectens |
 | Table 12- Summary of qualitative and quantitative assays of digestive enzymes in the gut of P.annctens n=60 |