ISSN : 0975-5276
EISSN : 0975-9174

ELSHAFIE H.S.1, CAMELE I.2*, BUFO S.A.3
1School of Agricultural, Forestry, Food and Environmental Sciences, University of Basilicata, Viale dell’Ateneo Lucano, 10, 85100 Potenza, Italy.
2School of Agricultural, Forestry, Food and Environmental Sciences, University of Basilicata, Viale dell’Ateneo Lucano, 10, 85100 Potenza, Italy.
3Department of Science, University of Basilicata, Via dell’Ateneo Lucano, 10, 85100 Potenza, Italy.
* Corresponding Author : ippolito.camele@unibas.it
Received : 01-03-2013 Accepted : 28-03-2013 Published : 01-04-2013
Volume : 5 Issue : 3 Pages : 399 - 403
Int J Microbiol Res 5.3 (2013):399-403
DOI : http://dx.doi.org/10.9735/0975-5276.5.3.399-403
Numerous species in the genus Burkholderia have interesting properties for potential industrial applications including production of antibiotics, biosurfactants, bioplastics and degradation of environmental contaminants. The aim of this research was study the effect of different carbon and nitrogen sources on Burkholderia gladioli pv. agaricicola strain ICMP11096 growth and bioactivity of produced secondary metabolites against gram-positive bacterium Bacillus megaterium and plant pathogenic fungi Rhizoctonia solani. The additional carbon sources were sucrose, fructose and lactose whereas, the additional nitrogen sources were, urea, potassium nitrate and ammonium nitrate. The results demonstrated that the addition of new carbon and nitrogen sources to the minimal mineral nutrient media lead to decreasing the growth rate of studied bacterial strain and increasing the production of bioactive substances. The maximum production of bioactive substances of studied bacterial strain was obtained using ammonium nitrate and lactose. The purified filtrate of the studied bacterial strain was fractionated by High Performance Liquid Chromatography (HPLC) and the antimicrobial activity of five isolated single peaks was evaluated against gram positive bacteria B. megaterium ITM100 and gram negative bacteria Escherichia coli ITM103. The most bioactive peak was number two with 12800 and 6400 Ua.ml-1 against B. megaterium and E. coli, respectively. The obtained results suggested that the nutrient amendments can increase the production of antimicrobial substances and this may be a useful strategy for improving the biocontrol efficiency.
Antimicrobial activity, Secondary metabolites, Carbon sources, Nitrogen sources, Bacterial growth rate.
Increasing the environmental concerns of production the bioactive microbial substances for application as biopesticides are considered promising alternatives to synthetic pesticides. There are some important factors influencing the bacterial growth including physical requirements and available sources of chemical nutrients [1] .
The production of bacterial secondary metabolite depend, also, on the nutrient status of culture medium [2,3] and is related with some various aspects of gene regulations and environmental factors [4,5] . The minimal mineral agar (MMA) is considered the best nutrient media suitable for study the growth of Burkholderia sp. [6] because it contains the minimum nutrients possible for bacterial growth. A defined medium has known quantities of all ingredients to provide a defined carbon and nitrogen source as well as essential trace elements and vitamins required by the microbe [7] . Carbon is the structural backbone of the organic compounds that make up a living cell [8] such as: glucose, galactose, sucrose and starch. Nitrogen is needed for the synthesis of such molecules as amino acids, DNA, RNA and ATP [5] .
Demain [9] , observed that the biosynthesis of the microbial secondary metabolites is induced by the addition of a nutrition system and by decreasing the growth rate so that the bacterial cells could reproduce slowly and reach to the maximum colony form unit (CFU) in longer period and the bioactivity of produced metabolites increased gradually to reach the maximum value during the stationary phase.
N.Acyl Homoserine lactones (N.AHLs) signal molecule based quorum sensing (QS) phenomenon play a vital regulatory role in chemical differentiation of secondary metabolism and morphological differentiation of morphogenesis which is directly proportional with the CFU [9-12] . Recent investigations have shown that Burkholderia gladioli pv. agaricicola (Bga) produces the above signal molecule [13] and have confirmed the production of group secondary metabolites, which could include novel active compounds [14] . Some strains of Bga have showed antifungal activity against some serious phytopathogenic fungi [15] . The aims of the present research were: (i) study the effect of different carbon and nitrogen sources on Bga ICMP11096 growth and increasing the production of its bioactive secondary metabolites, (ii) study the bioactivity of produced metabolites in modified substrates against gram-positive bacterium Bacillus megaterium (B.meg) and plant pathogenic fungi Rhizoctonia solani (R.sol), (iii) HPLC fractionation of produced secondary metabolites of Bga ICMP11096 and evaluate the bioactivity of isolated peaks against B.meg and Escherichia coli ITM103 (E.col).
The wild type strain of Bga ICMP11096 was isolated from Agaricus bitorquis (Quelet) Saccardo and obtained from ICMP Collection of Landcare Research (Auckland, New Zealand). This strain was kept as pure-freeze lyophilized stock maintained collection of School of Agricultural, Forestry, Food and Environmental Sciences, Basilicata University (Potenza, Italy). The studied target phytopathogenic fungus was R.sol whereas the target bacterial strain was gram positive B.meg ITM100 and E.col ITM103. The tested fungi and bacteria strains were derived from a pure culture maintained collection of the same department kept on potato dextrose agar (PDA) and King B (KB) respectively at 8°C.
Three different carbon and nitrogen sources were supplemented separately to the MM liquid broth media as following; sucrose 5 g.l-1, fructose 5.2 g.l-1 and lactose 5.2 g.l-1 are containing 0.175 mols of carbon atoms and urea 3.0 g.l-1, potassium nitrate 10.1 g.l-1 and ammonium nitrate 4.0 g.l-1 are containing 0.1 mols nitrogen atoms. Erlenmeyer flasks containing 25ml of nutrient MM broth media were inoculated with 1 ml suspension of Bga ICMP11096 containing about 108 CFU.ml-1 (optical density, O.D. 0.2 at 590 nm) from fresh culture of 24h age, and were incubated at 25°C under shaking (180 rpm). Three replicates were prepared for each nutrient treatment. The additional cultures were prepared as control composed only from MM plus each carbon and nitrogen sources.
The preparation of bacterial filtrates was carried out by centrifugation of broth culture at 20.000 g for 15 min and filtration using sterile Millipore filter 0.22 mm. For the antifungal test, R.sol was grown on PDA medium and agar well diffusion method was carried out using the method of Magaldi et al. [16] as following: wells of 0.5 cm diameter were punched into the nutrient medium and filled with 10 µl of prepared cell-free culture filtrates using serial dilutions (1:1 v/v). The plates were left for 24 h in a refrigerator at 4°C for completely diffusion. The plates were then incubated at 25°C for 96h. In case of antibacterial test, B.meg was grown on KB medium and the test was carried out using the method of Lavermicocca et al. [17] as following: 10 µl using serial dilutions (1:1 v/v) of cell-free culture filtrates were dropped on 10 ml of solid KB (in a 90x10 mm Petri dish) [18] . Completely dryness, 4 ml of soft agar (0.7 %) containing 0.5 ml of bacterial target suspension of 108 CFU.ml-1 of B.meg was poured on the plate surface and then the plates were incubated at 25°C for 48h. The activities were expressed as Unit active for millilitre (Ua.ml-1) where unit is the amount of substance contained in 10 ml capable of inhibiting the growth of the target organism [19] .
The bacterial growth was measured for each sample using spectrophotometer UNICAM-UV 500 apparatus at 600 nm to determine the optical density that refer to the bacterial growth in each media.
The purified filtrate of the most bioactive Bga strain ICMP11096 was analysed by High Performance Liquid Chromatography (HPLC). The analysis involved two main stages: sample preparation and fractionation by HPLC.
Aliquots (30 mg.ml-1) of lyophilized bioactive 5 days old culture filtrates of wild type Bga strain ICMP11096 was taken up in solution in sterile distilled water and the resulting solution loaded on cartridge syringe (Strata C18-T) prewashed with 2 ml methanol followed by 2 ml distilled water. The loaded solution was 10 ml containing 300 mg of lyophilized substances. After that, the cartridge was washed with 1 ml distilled water and recovered by 1 ml of methanol that was collected in sterilized eppendorf.
The methanol fraction was analyzed in the analytical HPLC-Agilent 1200 series austere. The best separation was obtained with an Agilent ECLIPSE XDB, C18 (4.6 x 150 mm, 5 μm) in analytical conditions. The injected volume was 20 μl, column temperature was 25°C, autosampler chamber temperature was 4°C, flow rate was s 1.0 ml.min-1 and mobile phase were: A: 0.2% Formic acid (FA) in H2O and B: 0.1% FA in Acetonitrile (CH3CN). The HPLC chromatogram was obtained at wave length l= 380 nm. The antimicrobial activity of isolated single peaks was evaluated against B.meg ITM100 and E.col ITM103 as reported before by Lavermicocca et al., [17] and the bioactivity was expressed as Ua.ml-1 [19] .
The maximum antifungal activity against R.sol was observed using ammonium nitrate with lactose and fructose in nutrient media. The activity was 3200 Ua.ml-1 after 13 days of incubation [Table-1] . The increase of bioactive substances was observed between 3-7 days using ammonium nitrate with fructose and lactose and reached a maximum after 13 days compared to control. These results are considered the positive effect for using ammonium nitrate as nitrogen source with all studied carbon sources.
The maximum antibacterial activity against B.meg was observed using ammonium nitrate with lactose and fructose. The activity was 6400 Ua.ml-1 after 13 days of incubation [Table-2] . The increase of bioactive substances was observed between 3-7 days and reached a maximum after 13 days compared to control. The obtained results of antimicrobial activity confirmed the hypothesis that the addition of carbon sources in the beginning of life cycle of bacteria plays an essential role in prolonging the period of production of the bioactive substances due to the best adaptation of tested bacteria to the nutrient media during the log phase than stationary phase and hence the net production of the total active substances were maximized at the end of stationary phase in accordance with Hengge-Aronis and Lazazzera [20,21] who reported that the transition into stationary phase is directed by some key transcription factors which are stimulated by starvation.
The mid log phase in control media started faster than using urea with all tested carbon sources where the maximum growth achieved after 9 days of incubation [Fig-1] . Using potassium nitrate leads to prolong the log phase compared to control media especially with lactose and fructose due to the reduction of growth rate and the maximum growth was achieved after 13 days in case of potassium nitrate with fructose [Fig-2] . Using ammonium nitrate leads to prolonging the log phase compared to control media indicating the highly adaptation of bacteria toward the new nutrient components especially with lactose, fructose and sucrose respectively. The maximum growth using ammonium nitrate and lactose was achieved after 13 days [Fig-3] .
The obtained results of optimization the nutrient media could investigate the vital regulatory role of N.AHLs signal molecule in the production of secondary metabolites. This signal molecule can be produced with high amount at the maximum bacterial cell density to induce the high production of secondary metabolites with maximum bioactivity [22,23] . The nutrient media components effect on the expression of quorum sensing phenomenon which plays a central role in the gene regulation of specific bacterium to encode the more efficient functions at high cell density such as production of secondary metabolites [24,25] . The above hypothesis is confirmed by the low molecular weight nature of the signal molecules for inactivating the regulatory protein to decrease the production of secondary metabolites and enhance the morphogenesis during the rapid growth rate. So the decrease of bacterial growth rate induces the biosynthesis of secondary metabolites and reaches to the maximum production in parallel with the formation of signal molecules at high cell density in agreement with the results obtained by Demain [9] .
From chromatographic runs conducted at l = 380 nm, it was noticed the presence of five single peaks [Fig-4] . The five peaks were collected and evaluated for their antimicrobial activity against B.meg and E.col. In particular; the most bioactive peaks were number 2 and 3 [Table-3] , [Fig-5] . The identification of bioactive peaks is in progress.
Results obtained in this study suggested that the nutrient amendments may increase the production of antimicrobial substances and this may be a useful strategy for improving the biocontrol efficiency in accordance with Duffy and Defago [26] . Moreover, the addition of new carbon and nitrogen sources at the beginning of lag phase is useful to prolong the transition into stationary phase due to decreasing the competition between bacterial colonies and lowering the growth rate of bacterial cells which resulting in highly production of bioactive substances as well. On the other hand, the addition of those amendments during the mid log phase causes a highly competition and increases the growth rate and decreasing the production of antimicrobial substances. In fact, the bacterial colonies must adapted themselves to the nutrient media components during the lag phase for inducing the specific genes for production the antimicrobial substances and reach to the maximum growth at the end of exponential phase in order to enter the stationary phase and produce the highest quantity of bioactive substances [27,28] . Generally, in the present study, it was concluded that the maximum antimicrobial activity of Bga ICMP11096 was obtained after incubation for 13 days at 25°C in MM nutrient media adjusted at 7.0 pH and supplemented with ammonium nitrate 4 g.l-1 and lactose 5.2 g.l -1.
We are grateful to Dr. Parisien Michèle Lynn Marie (University Language Centre-UNIBAS-CLA) for English revision and thanks for the technical assistance from C.T. Michele Palumbo.
The authors declare no conflict of interest.
Bacillus megaterium : B.meg
Burkholderia gladioli pv. agaricicola : Bga
Colony form unit : CFU
Escherichia coli : E.col
High Performance Liquid Chromatography : HPLC
International Collection of Microorganisms from Plants : ICMP
King B : KB
Minimal Mineral Agar : MMA
N.Acyl Homoserine lactones : N.AHLs
Potato Dextrose Agar : PDA
Quorum sensing : QS
Rhizoctonia solani : R.sol
Unit active of millilitre : Ua.ml-1
[1] Prapagdee B., Kuekulvong C. and Mongkolsuk S. (2008) Int. J. Biol. Sci., 4 (5), 330-337.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[2] El-Banna N. and Winkelmann G. (1998) J. Appl. Microbiol., 85, 69-78.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[3] Hwang J., Chilton W.S. and Benson D.M. (2002) Biol. Control, 25, 56-63.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[4] Duffy B.K. and Defago G. (2000) Appl. Environ. Microbiol., 66, 3142-3150.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[5] Young S.K., Lee Y.J., Lee Y.H. and Kim J.H. (2009) J. Microbiol. Biotechnol., 19(10), 1142-1149.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[6] Howard K. and Inglis T.J. (2005) Water Res., 39(6), 1085-1092.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[7] Madigan M. and Martinko J. (2005) Brock Biology of Microorganisms. San Francisco, CA: Benjamin.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[8] Fagerbakke K.M., Heldal M. and Norland S. (1996) Aqatic Microb. Ecol., 10, 15-27.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[9] Demain A.L. (1998) Int. Microbiol., 1, 259-264.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[10] Withers H., Swift S. and Williams P. (2004) Curr. Opin. Microbiol., 4, 186-193.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[11] Liu X., Bimerew M., Ma Y., Muller H., Ovardis M., Eberl L., Berg G. and Chernin L. (2007) FEMS Microbiol. Lett., 270, 299-305.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[12] Duerkrop B.A., Ulrich R.L. and Greenberg E.P. (2007) J. Bacteriol., 189, 5034-5040.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[13] Prashanth S.N., Bianco G., Cataldi T.R. and Iacobellis N.S. (2011) J. Agric. Food Chem., 59, 11461–11472.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[14] Lamorte D., Elshafie H.S., Lelario F., Bufo S.A. and Iacobellis N.S. (2010) 12th Symposium on Chemistry and Fate of Modern Pesticides, Matera- Italy. 350-351.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[15] Elshafie H.S., Camele I., Racioppi R., Scrano L., Iacobellis N.S. and Bufo S.A. (2012) Int. J. Mol. Sci. 13, 16291-16302.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[16] Magaldi S., Mata-Essayag S., Hartung de Capriles C., Perez C., Colella M.T., Olaizola C. and Ontiveros Y. (2004) Int. J. Infect. Dis., 8, 39-45.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[17] Lavermicocca P., Iacobellis N.S., Simmaco M. and Graniti A. (1997) Physiol. Mol. Plant Pathol., 50, 129-140.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[18] King E.O., Ward M.K. and Raney D.E. (1954) J. Lab. Clin. Med., 44, 301-307.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[19] Sinden S.L., DeVay J.E. and Backman P.A. (1971) Physiol. Plant Pathol., 1, 199-213.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[20] Hengge-Aronis R. (1999) Curr. Opin. Microbiol., 2, 158-152.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[21] Lazazzera B.A. (2000) Curr. Opin. Microbiol., 3, 177-182.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[22] Lazazzera B.A., Palmer T., Quisel J. and Grossman A.D. (1999) Cell-Cell Communication in Bacteria, ASM Press, 27-46.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[23] Seo Y.S., Lim J., Choi B.S., Kim H., Goo E., Lee B., Lim J.S., Choi I.Y., Moon J.S., Kim J. and Hwang I. (2011) J. Bacteriol., 193, 3149-3149.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[24] Velusamy P. and Gnanamanickam S.S. (2008) Secondary Metabolites in Soil Ecology, Springer-Verlag Berlin Heidelberg, 5.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[25] Elshafie H.S., Iacobellis N.S. and Bufo S.A. (2012) Fifth Saudi Science Conference (SSC5), 92-93.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[26] Duffy B.K. and Defago G. (1999) Appl. Environ. Microbiol., 65, 2429-2438.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[27] You Z., Fukushima J., Tanaka K., Kawamoto S. and Okuda K. (1998) FEMS Microbiol. Lett., 164, 99-106.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[28] Steindler L. and Venturi V. (2007) FEMS Microbiol. Lett., 266, 1-9.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
 | Fig. 1- Bacterial growth curve of Burkholderia gladioli pv. agaricicola ICMP11096 with N1 (C1, C2, C3). C1: Sucrose, C2: Lactose, C3: Fructose and N1: Urea. |
 | Fig. 2- Bacterial growth curve of Burkholderia gladioli pv. agaricicola ICMP11096 with N2 (C1, C2, C3). C1: Sucrose, C2: Lactose, C3: Fructose and N2: Potassium nitrate. |
 | Fig. 3- Bacterial growth curve of Burkholderia gladioli pv. agaricicola ICMP11096 with N3 (C1, C2, C3). C1: Sucrose, C2: Lactose, C3: Fructose and N3: Ammonium nitrate. |
 | Fig. 4- HPLC chromatogram of ICMP11096 of Burkolderia gladioli pv. agaricicola at 380 nm. |
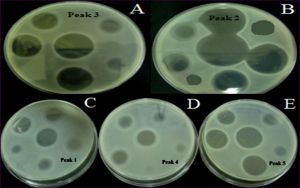 | Fig. 5- Antimicrobial activity of single HPLC peaks vs. Bacillus megaterium. Where: A: Peak 3, B: Peak 2, C: Peak 1, D: Peak 4 and E: Peak 5. |
 | Table 1- Antifungal activity of Burkholderia gladioli pv. agaricicola strain ICMP11096 against Rhizoctonia solani. C1: Sucrose, C2: Lactose, C3: Fructose, N1: Urea, N2: Potassium nitrate and N3: Ammonium nitrate. |
 | Table 2- Antibacterial activity of Burkholderia gladioli pv. agaricicola strain ICMP11096 against Bacillus megaterium. C1: Sucrose, C2: Lactose, C3: Fructose, N1: Urea, N2: Potassium nitrate and N3: Ammonium nitrate. |
 | Table 3- Antimicrobial activity of single HPLC peaks |