ISSN : 0975-3702
EISSN : 0975-9182

RADWAN N.A.1*, ABOU SHAFEEY H.E.2, KHALIL A.I.3
1Department of Zoology, Faculty of Science, University of Tanta, Tanta, Egypt
2Department of Zoology, Faculty of Science, University of Tanta, Tanta, Egypt
3Department of Zoology, Faculty of Science, University of Tanta, Tanta, Egypt
* Corresponding Author : nahla_ahmad_mohamed@yahoo.com
Received : 10-11-2012 Accepted : 03-12-2012 Published : 18-12-2012
Volume : 4 Issue : 2 Pages : 100 - 105
Int J Parasitol Res 4.2 (2012):100-105
DOI : http://dx.doi.org/10.9735/0975-3702.4.2.100-105
Conflict of Interest : None declared
The tegumental spines around the attachment organs in four species of digenetic trematodes namely; Fasciola gigantica, Schistosoma heamatobium, Cynodiplostomum azimi and Aporchis massiliensis were evaluated with energy dispersive X-ray microanalysis (EDXA) for three chemical elements: sulphur, calcium and phosphorus at three regions of spine (tip, middle and base) and the average was recorded. Weight percent (wt %) of each chemical element for each digenean at different regions was recorded and analyzed for probability using SPSS computer program. The pattern of element distribution showed significant specific characteristics. In F. gigantica (distome type), sulphur had the highest concentration followed by calcium then phosphorous, in Aporchis massiliensis (echinostome type), calcium concentration was the highest followed by sulphur while phosphorous was completely absent. In S. heamatobium (shistosome type) and C. azimi (holostome type), which follow the same pattern, phosphorous had the highest concentration followed by sulphur then calcium. The functional significance of the variations in element distribution in different parts of the spines in diverse digenean types is most likely related to the habitat richness with these elements in addition to the ability of these digenean types to metabolize and make use of these elements.
Fasciola gigantica, Schistosoma heamatobium, Cynodiplostomum azimi, Aporchis massiliensis, EDXA, tegument.
Energy dispersive analysis for X- ray (EDXA) is a non- destructive technique whereby the sample can be analyzed several times at different locations and magnifications [1,2] . X- ray microanalysis has been used successfully in many research disciplines. Parasitic diseases have been further understood by using this technique [3] , where evidence for many pathological problems has employed EDXA [4] . In the parasitological field, EDXA has been used for analysis of chemical elements in the body of helminthes, specially the anchorage structures [5,6] . Studies used the variation in the concentrations and distribution of chemical elements to specify and correlated these variations with type, type host and habitats [7-9] . This technique was also employed to determine the effects of exponential growth of some helminthes on the concentration of some metabolically active elements inside the hosts [10] .
Spines are anchorage structures present in many digenean species. The chemical and structural characteristics may help in understanding its functional role in different digenean groups.
The goal of this study is to evaluate three chemical elements: sulphur (S), calcium (Ca) and phosphorous (P) in the spines around the attachment organs (sucker) of four morphological types of digenetic trematodes and correlate the concentration and distribution of these elements to the possible anchorage function of each type. Data is evaluated statistically using SPSS program (ANOVA).
Adult Fasciola gigantica, Schistosoma heamatobium, Cynodiplostomum azimi and Aporchis massiliensis were collected from the bile duct of cattle, blood of albino mice, intestine of mice and intestine of white-eyed gull respectively. Specimens were fixed in 70% ethyl alcohol and dehydrated in ascending series of ethyl alcohol. Standard methods were followed to prepare each specimen for SEM and X-ray microanalysis [11] which included critical point drying (CPD) and mounting on SEM sample mounts using conductive double sided carbon tape. Mounted specimens were gold coated for 30 seconds (approximate gold coating thickness of 20nm). Samples were examined in a JOELJXA-840A electron microscope in combination with INCA X-sight X ray analyzer. Samples were imaged at 30KV. EDXA scans were completed for triplicate specimens of the spines around the attachment organ (ventral sucker of Fasciola gigantica and Schistosoma heamatobium; oral sucker of Aporchis massiliensis and pseudosuckers of Cynodiplostomum azimi) at three regions (tip, middle and base) and the average was recorded. Weight percent (wt %) of each chemical element for each digenean at different regions was recorded [Fig-1] and analyzed for probability using SPSS computer program.
The concentration of three chemical elements, namely sulphur, calcium and phosphorus in the tegumental spines around the ventral suckers of Fasciola gigantica and Schistosoma heamatobium, pseudosuckers of Cynodiplostomum azimi and oral sucker of Aporchis massiliensis [Fig-2a] , [Fig-2b] , [Fig-2c] , [Fig-2d] are demonstrated in [Table-1] and [Fig-3] .
In the spine of Fasciola gigantica, S had the highest concentration followed by Ca then P. In Aporchis massiliensis, Ca was the element of highest concentration followed by S, while P was absent. In both Schistosoma heamatobium and Cynodiplostomum azimi, P was the element of highest concentration followed by S while Ca had the lowest concentration.
Highest concentration of S was detected in Fasciola gigantica followed by Cynodiplostomum azimi, then Schistosoma heamatobium and the lowest concentration was detected in Aporchis massiliensis.
Fasciola gigantica had the highest concentration of Ca followed by Aporchis massiliensis then Cynodiplostomum azimi and the lowest concentration was detected in Schistosoma heamatobium.
P demonstrated the highest concentration in Schistosoma heamatobium followed by Cynodiplostomum azimi then Fasciola gigantica, while no P was detected in Aporchis massiliensis.
One way ANOVA [Table-2a] , [Table-2b] and [Table-2c] revealed high significant differences in the concentration of the three elements between the four species under study.
The level of sulphur, calcium and phosphorous was determined for spines around the attachment organ of the four digeneans in three regions of the spine, namely the tip, middle and base [Fig-4] . [Table-3] shows that the distribution of S throughout the spine varied in the four examined species. In both Fasciola gigantica and Aporchis massiliensis, the concentration of S gradually increased from the tip till the middle region of the spine, then decreased towards the base where it reached its lowest concentration. However, the tip had a higher concentration than the base. In both Schistosoma heamatobium and Cynodiplostomum azimi the concentration of S was high at the tip of the spine and decreased towards the middle region then increased again posteriorly towards the base of the spine. Moreover, the concentration of this element in the spine base was higher than its concentration at the spine tip.
The concentration of both Ca and P follow the same pattern, where in both Fasciola gigantica and Aporchis massiliensis, both elements reached their highest levels at the tip of the spine and gradually decreased posteriorly towards the base region where lowest concentrations were recorded. No Ca and P were detected in the spine base of Aporchis massiliensis. In both Schistosoma heamatobium and Cynodiplostomum azimi, the concentration of both elements was high at the spine tip and decrease towards the middle region of the spine, then increase again towards the base region. Likewise, their concentration at the base of the spine was higher than that at the spine tip.
One way ANOVA was performed to detect differences in the distribution of the three elements in the different spine regions in the same [Table-4] as well as between the four examined species [Table-5] , [Table-6a] , [Table-6b] , [Table-6c] . [Table-4] shows that there is a significant difference in the concentration of both Ca and P throughout the spine regions in Fasciola gigantica, S and Ca in Aporchis massiliensis and S and P in Schistosoma heamatobium. No significant difference was detected in the concentrations of the three elements within the different regions of the spines of Cynodiplostomum [Table-5] [Table-6a] , [Table-6b] and [Table-6c] show significant differences in the three chemical elements versus the there regions of the spine in the four examined species.
In the tip and the middle regions of the spine, the concentration of S, Ca and P showed a significant difference between Fasciola gigantica and each of Aporchis massiliensis, Schistosoma heamatobium and Cynodiplostomum azimi.
The base region of the spine showed a significant difference in the concentration of S between Fasciola gigantica and each of Aporchis massiliensis and Schistosoma heamatobium, also between Aporchis massiliensis and Schistosoma heamatobium and Cynodiplostomum azimi and between Cynodiplostomum azimi and Schistosoma heamatobium. As regards Ca and P concentration, there were significant differences between Fasciola gigantica and each of Schistosoma heamatobium and Cynodiplostomum azimi, and between Aporchis massiliensis and both Schistosoma heamatobium and Cynodiplostomum azimi.
Trematodes have adapted in different ways to their microenvironment within the vertebrate host. Hayunga [12] demonstrated that life in this specialized habitat presents special challenges of exposure to digestive enzymes, normal peristalsis, and host immune response to infection and that the surface of the parasite should be the first part of the organism to encounter such challenges. Morphological adaptations of trematodes to their microenvironment include modification of the tegumental surface that includes the development of specialized attachment organs, such as the suckers. The external surface of the tegument is associated with unique ridges among which are dispersed spines that varies structurally in different species. The shape, distribution, orientation and composition of the spines especially in the tegument around the suckers help in understanding the anchorage mechanisms of such structures and their role in life enhancement of trematodes.
Sulphur, calcium and phosphorous are major elements in biological molecules of all representatives of the animal kingdom. Furthermore, they are associated with the development and function of helminthes hooks and spines [7-9] . Phosphorous aids in the formation of bones and teeth along with calcium, it gives the hardness, strength and rigidity to resist compressive forces. Carbon and sulphur are main constituents of keratin which is the building component of hair, horns, hooves, beaks, shells, nails, claws … etc.
Gallagher [5] and Smith and Richards [6] studied the rostellar hooks of cestodes referred to similarities with vertebrate keratin and discussed the validity of using the term keratin to refer to cestodes hook material.
The detection of the presence and differential distribution of these elements within the tegumental spines around the attachment organs of digeneans inhabiting different habitats would add to our understanding of the vital role of the spines as anchorage structures in life support of these parasites. This understanding will provide additional targets that can be exploid by chemotherapeutic means. In addition and because these elements are present in different concentrations in diverse helminthes, such information might be helpful in understanding the relationship between digenean species by revealing the intra and inter-specific degree of similarity and divergence of the chemical and physical composition of these structures.
The present study was conducted to evaluate the levels of three chemical elements: sulphur, calcium and phosphorus, using energy dispersive X-ray analysis (EDXA), in the different spine regions of four digenean species related to four morphological groups inhabiting different habitats.
The pattern of element distribution showed significant specific characteristics. In F. gigantica (distome type), sulphur had the highest concentration followed by calcium then phosphorous, while in Aporchis massiliensis (echinostome type), calcium concentration was the highest followed by sulphur while phosphorous was completely absent. In S. heamatobium (shistosome type) and C. azimi (holostome type), which follow the same pattern, phosphorous had the highest concentration followed by sulphur then calcium.
The large size of F. gingatica that lives in narrow hostile bile duct and C. azimi inhabiting the intestinal microhabitat which undergo peristaltic contraction need both S and Ca to provide hardness to the spines. On the other hand, the mobile coupled S. heamatobium needs mobility rather than anchorage support, thus restrict the need for hard spines.
The concentration of phosphorous was highest in the spines of S. heamatobium followed by C. azimi then F. gigantica. The colar spines of Aporchis massiliensis were free of phosphorous. The high concentration of P in Shistosoma might be related to its habitat «The blood».
In the collar spine of Aporchis massiliensis, the very low concentration of sulphur with high concentration of calcium and absence of phosphorous is unexplainable. However the high calcium concentration fits well with the report of Klenchin et al. [13] who referred to the presence of Ca+2 binding sites in actin monomers, which are the subunits of the microfilaments and one of the three major components of all cytoskeletons, in the collar spines of Echinostoma trivolvis [14] . The collar spines of adult echinostomatids function mainly in worm attachment to the host mucosa in addition to secretion [15] .
Differential distributions of the three elements throughout the spines revealed that F. gigantica and Aporchis massiliensis follow the same pattern and S. heamatobium and C. azimi share the same pattern. In both F. gigantica and Aporchis massiliensis, sulphur increase from the tip of spine towards the middle regions where it reaches its highest levels then decrease again towards the base. On the other hand, sulphur in both S. heamatobium and C. azimi has a high concentration on the tip and decrease towards the middle region, where it has its lowest values then it increase again and reaches its highest concentration at the base. However the concentration of sulphur was high in the base than the tip.
Calcium and phosphorous had the same distribution pattern, in both F. gigantica and Aporchis massiliensis, both elements reached their highest concentration at the tip and gradually decreased throughout the spine till the base where the lowest concentrations were recorded. While in S. Heamatobium and C. azimi the concentration of both elements decrease towards the middle region then increase again to reach their highest levels at the base.
Heckmann et al. [9] clarified that the calcium and phosphorous in the acanthocephalan proboscis hooks form a rigid calcium and phosphorous complex similar to the enamel of mammalian teeth with disulphide bonds (cysteine) enhancing the structure. The authors predicted that the less sulphur at the base of the proboscis hook creat a more flexible area for the hook, while the high sulphur content at the tip of the hook and along the edges is probably polymerized into a complex protein with disulfide bonds. They referred to the important role of disulfide bonds in the stability of the protein which is formed between the thiol groups of the amino acids and added that cystiene and cystine are probably the main sulphur containing amino acids for the hooks of Acanthocephala.
The functional significance of the variation in element distribution in different parts of the spines in diverse digenean types is hard to understand, but it is most likely related to the habitat richness with these elements in addition to the ability of these digenean types to metabolize and make use of these elements. The results revealed clearly that high calcium and phosphorous concentration in the spine tip of F. gigantica and Aporchis massiliensis makes for the low sulphur and might represent components of calcium phosphate that give the spine rigidity, the high sulphur concentration in the spine body give rigidity and make for the decreasing calcium and phosphorous content. The base is low and free of calcium and phosphorous in Aporchis massiliensis is unexplainable, except if this low elements concentration allows the spine flexibility to fit in the host parasite interface.
Whether these variations are species specific or individual variations, necessitates evaluation of the element composition of a larger number of species inhabiting different habitats, as well as studying spines in different parts of the body and in different stages of development of parasites.
[1] Sures B. (2003) Parasitology, 126, 53-60.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[2] Whallon J.H., Flegler S.L., Klomparens K.L. (1989) Bioscience 39, 256-259.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[3] Heckmann R.A. (1997) Microscopy and Analysis, 18, 35-37.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[4] Ericksen M. (1997) Light and Electron Microscopy Including X-Ray Microanalysis of Human Prostate Concentrations in Relation to Prostate Cancer. MS Thesis. Brigham Young University, Provo, Utah, 34.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[5] Galagher H.C. (1964) Exp. Parasitol., 15, 1100-1117.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[6] Smith S.A., Richards K.S. (1991) Parasitology, 103, 267-274.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[7] Heckmann R.A., Amin O.M., Standing M.D. (2007) Comp. Parasitol., 74, 388-391.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[8] Heckmann R.A., Amin O.M., Radwan N.A., Standing M.D., Eggett D.L. (2012)Sci. Parasitol., 13(1), 27-35.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[9] Heckmann R.A., Amin O.M., Radwan N.A., Standing M.D., Eggett D.L., El Naggar A.M. (2012) Sci. Parasitol., 13(1), 37-43.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[10] Wranicz M., Dabrowska M., Zielinski Z., Michalski R., Pawelczak K., Rode W. (1996) Biochem Biophys Res Commun., 12, 288(2), 440-445.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[11] Lee P.E. (1992) Scanning Electron Microscopy and X-ray Microanalysis. Prentice Hall, Englewood Cliffs, New Jersey, 458.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[12] Hayunga E.G. (1991) J. Parasitol., 77(6), 865-73.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[13] Klenchin V.A., Khaitlina S.Y. and Rayment I. (2006) J. Mol. Biol., 362, 140-150.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[14] Fujino T., Fried B. and Hosier D.W. (1994) Parasitol. Res., 80(7), 581-587.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[15] Fried B. and Toledo R. (2009) Parasit. Vectors., 2, 52.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
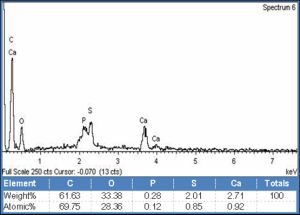 | Fig. 1- X-ray Microanalysis of the tip of the spine around the ventral sucker of Fasciola gigantica, showing chemical elements at their KeV peak with the wt % of each element. The chart represents an example of 36 charts covering all spines region around the attachment organ in the four encountered digeneans. |
 | Fig. 2a- Tegumental spines around ventral sucker of F. gigantica |
 | Fig. 2b- Tegumental spines around ventral sucker of Shistosoma heamatobium |
 | Fig. 2c- Tegumental spines around oral sucker of Aporchis massiliensis |
 | Fig. 2d- Tegumental spines around pseudo sucker of Cyndoplostomum azimi |
 | Fig. 3- Concentrations of Sulphur, calcium and phosphorus in the tegumental spines around the attachment organs in the four digeneans. |
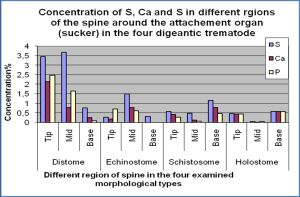 | Fig. 4- Concentrations of of Sulphur, calcium and phosphorus in different regions of the tegumental spines around the attachment organs in the four digeneans. |
 | Table 1- Weight percent (wt%) of sulphur (S), calcium (Ca), phosphorus (P) in the tegumental spine around the attachment organ of the four digenean species. |
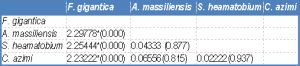 | Table 2a- Statistical analysis: Mean difference (P value) of EDXA data of sulphur in tegumental spine around attachment organ. |
 | Table 2b- Statistical analysis: Mean Difference (P value) of EDXA data of calcium in tegumental spine around the attachment organ. |
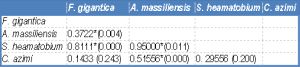 | Table-2c Statistical analysis: Mean Difference (P value) of EDXA data of phosphorus in tegumental spine around the attachment organ |
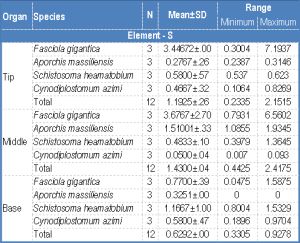 | Table 3- Weight percent (wt%) of sulphur (S), calcium (Ca), phosphorus (P) in the tip middle and base of the spine around the attachment organ of the four digenean species |
 | Table 4- Statistical analysis: P value of EDXA data of S, Ca and P in spine regions in the four encountered species |
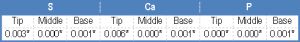 | Table 5- Statistical analysis: P value of EDXA data of S, Ca & P in tip, middle and base of spines in the different encountered species |
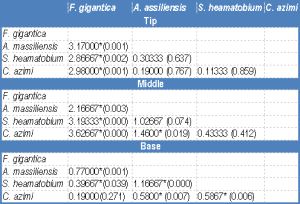 | Table 6a- Statistical analysis: mean difference (P value) of EDXA data of sulphur in three regions (tip, middle and base) of spine around the attachment organ |
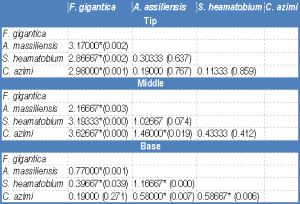 | Table 6b- Statistical analysis: mean difference (P value) of EDXA data of Calcium in three regions (tip, middle and base) of spine around the attachment organ |
 | Table 6c- Statistical analysis: mean difference (P value) of EDXA data of Phosphrus in three regions (tip, middle and base) of spine around the attachment organ |