ISSN : 0976-5530
EISSN : 0976-5549

ANANDHAGUNADEVI1, RAMANATHAN VADUKKUPATTU DEVASENAPATHI2, RATHINAVEL ANDIAPPAN3, SHAKILA HARSHAVARDHAN4*
1Department of Plant Biotechnology, School of Biotechnology, Madurai Kamaraj University, Madurai- 625021, India
2Department of Clinical pathology, Tuberculosis Research Centre, Chennai- 600 031, India
3Department of Cardio-Thoracic surgery, Government Rajaji Hospital , Madurai- 625 020, India
4Department of Plant Biotechnology, School of Biotechnology, Madurai Kamaraj University, Madurai- 625021, India
* Corresponding Author : mohanshakila@yahoo.com
Received : 09-09-2011 Accepted : 28-09-2011 Published : 01-12-2011
Volume : 2 Issue : 2 Pages : 34 - 40
Int J Med Clin Res 2.2 (2011):34-40
DOI : http://dx.doi.org/10.9735/0976-5530.2.2.34-40
Conflict of Interest : None declared
Acknowledgements/Funding : The authors acknowledge the patients and the healthy
volunteers who gave their blood sample for the above
analysis
Background: Rheumatic heart disease is an inflammatory disease which predominantly affects the valvular endocardium culminating in crippling valve deformities. Inorder to find out the role of complement proteins in inducing the pathogenesis, serum analysis of complement proteins C3a, C4a, C5a, C3c and C3d and acute phase protein MBL in patients with Rheumatic heart disease were performed. Methods and Results: Blood samples were collected from twenty eight subjects who were diagnosed with Rheumatic heart disease (RHD). Same number of samples was also collected from healthy volunteers as control. The levels of the anaphylatoxins namely C3a, C4a and C5a and the acute phase protein MBL (Mannose Binding Lectin) were assessed quantitatively in the samples using Sandwich ELISA. The breakdown products of the complement protein C3b namely C3c and C3d were assessed qualitatively using western blot assay. Among the three anaphylatoxins, the C5a level and the MBL level were significantly higher in RHD subjects than the control samples. The breakdown products of C3b namely C3c and C3d were observed only in RHD patients and not in control samples. Conclusions: Significant levels of C5a and MBL noticed in blood samples of RHD subjects when compared to the healthy controls indicate that these two proteins might corroborate the chronic inflammatory activity present in these individuals and contribute to valve injury through complement activation. Along with these results, the presence of C3c and C3d only in the RHD samples and not in the controls indicates that these products of C3 complement protein may also contribute for the inflammatory pathogenesis of the disease.
Rheumatic heart disease, anaphylatoxins, acute phase proteins, complement proteins and mannose binding lectin.
Rheumatic fever (RF) is a delayed sequelae to throat infection by Streptococcus pyogenes and affects susceptible untreated children. Based on the major criteria established by Jones and revised by the American Heart Association, the disease manifests as polyarthritis, carditis, chorea, erythema marginatum and â„ or subcutaneous nodules [1] . Nearly 75% of affected children display arthritis and 30–45% develop carditis, which causes heart damage with pericardial, myocardial and endocardial involvement followed by progressive and permanent valvular lesions leading to Rheumatic Heart Disease (RHD). It is usually a delayed manifestation and is often the sole manifestation of Acute Rheumatic Fever (ARF) [1] . RFâ„RHD is still a major public health burden in developing countries, leading to 233,000 deaths annually. The pathogenesis of RFâ„RHD is complex and both environmental and genetic factors contribute to its etiology. The manifestation of the disease in only a small subset of children untreated for strep throat and also the fact that only one-third of the affected children progress to the development of RHD suggests the involvement of host genetic factors [2] .
Complement system is not only vital for host defense against infectious diseases, but it also plays intricate roles in protection, development, and progression of inflammatory diseases including many rheumatic disorders [3] . As a key mediator of inflammation, its excessive or inappropriate activation through the classical, alternative or lectin pathways may contribute greatly to host tissue damage [3 & 4].
Thus, the present study was attempted to study the role of complement proteins in RHD. For this, the serum samples of the subjects diagnosed of RHD and the normal healthy individuals were analyzed quantitatively and qualitatively for the major anaphylatoxins C3a, C4a and C5a, which are the catabolic products of complement pathway. The Mannan Binding Lectin (MBL), an acute phase protein, involved in lectin pathway was also analyzed in these patients. Since RHD is an inflammatory disease of heart tissues, whether these proteins may be one of the initiation factors of the disease is imperative. The complement products of C3b i.e., C3c and C3d were also analyzed in both the above groups for they may also reveal the role of these proteins in this disease.
The subjects who were diagnosed with chronic RHD and were admitted in Rajaji hospital, Madurai, Tamilnadu, India, were enrolled for this study. Only twenty eight subjects were selected for this study before they had any treatment and hence blood samples were collected from them with their consent. For collecting the blood samples from the subjects and the healthy volunteers, prior approval was obtained from the Internal Research and Review Board (IRB), Ethical clearance (EC), Biosafety and Animal Welfare Committee, Madurai Kamaraj University, Madurai, Tamilnadu, India and also from Rajaji Hospital Ethical clearance Board, Madurai, Tamilnadu, India.
The subjects who had a clinical history of RF and an echocardiogram confirming mitral valve involvement were included in this study. Out of twenty eight patients who were enrolled for this study, 15 were males and 13 were females in the age group of 21-40 years. For control samples, blood was collected from 28 healthy volunteers out of which, 14 were males and 14 were females in the age group of 21-35 years.
Around 8ml to 10ml of venous blood sample was collected from the patients. Soon after collecting the blood samples, it was allowed to clot for 4hrs and centrifuged at 4oC for 10minutes at 1000xg. After centrifugation, the serum was collected from the blood samples and labeled. They were then distributed in aliquots and stored at -80oC until used.
The serum samples which were clear, non-hemolyzed and non lipemic were only used for this study. Samples were diluted prior to running the assay. An initial dilution of 1:500 of human serum of both patients and control samples with standard or sample diluents was done. All specimen handling operations was carried out at 4oC since the complement proteins are heat labile proteins.
The levels of serum C3a, C4a, C5a proteins (anaphylatoxins) were estimated using BD Opt EIaTM ELISA [Enzyme Linked Immunosorbant Assay] kit. The procedure was followed as per the instructions given in the kit. The kit utilizes monoclonal antibody (MAb) specific for human anaphylatoxin-des Arg coated on a 96 well plate. Standards and appropriately diluted samples were added to the wells and incubated at different intervals as mentioned in the kit. The wells were washed with wash concentrate (concentrated detergent solution with ProclinTM-150) and later a mixture of biotinylated polyclonal anti-human C3a, C4a or C5a antibody conjugated to streptavidin horseradish peroxidase was added producing Ab-Ag-Ab “sandwichâ€. The wells were again washed with wash concentrate and TMB (3, 3’, 5’, 5 Tetramethylbenzidine) substrate was added. The reaction was stopped with stop solution (1M H3PO4). The stop solution changes the color from blue to yellow and at this stage the wells were read in ELISA reader (BioRad) at 450nm within 30 minutes. All the standards and samples were run in duplicate. A standard curve was required in each assay run.
MBL oligomer ELISA kit (BD Biosciences) was used to measure the MBL oligomer level in the human serum. As per the instructions given in the kit, the serum of subjects and healthy people were diluted as 1:100 dilutions. Wells were precoated with MBL antibody. Similar methodology like the above sandwich assay was followed for measuring MBL.
Samples to be run in 10% poly acrylamide gel electrophoresis (PAGE) were first denatured in sample buffer by heating at 95-1000C for 5 minutes. Gel set up was done and the samples with higher titre values for C3a in ELISA were loaded along with control and marker in separate wells using syringe. The complete PAGE set up was connected to a power pack and 30mA volt of current was passed through the gel. Electrophoresis was ceased by tracking the BPB (Bromophenol blue) along the length of run and the gel was dismantled from the gel apparatus, discarded the stacking gel and placed the separating gel in staining solution. Staining was carried out with shaking for a minimum of 2hr then the stain was replaced with destaining solution.
Western Blot was done to qualitatively study the presence of C3c and C3d in serum using primary antibody of both polyclonal rabbit anti-human C3c complement and polyclonal rabbit anti-human C3d complement [1:500 dilution] [Dak] Denmark A/S] and secondary antibody of Eviflour fort orange 600 Goat F(ab’)2 Anti-rabbit IgG (H+L) (EvifloursTM-Quantum Dot Conjugates) was prepared in 1:100 dilution. Eviflour Quantum Dots are a ready to use flourescent probe for Western Blotting. Eviflours have semi-conductor nanocrystals at their core, which exhibit unique flourescent lifetime. These Eviflours are pre-conjugated to commonly used secondary antibodies, which subsequently can be used to target primary antibodies. Imaging of the western blot can be obtained using simple UV Epi Illumination Imaging systems (BioRad) with readily obtainable filters. In our study, we have utilized Biorad-gel analysis software, a Fluorescence Imaging System to photograph the fluorescent bands.
Data are given as mean ± SEM. Statistical differences between groups were determined by the two-tailed Student t -test. Differences were considered to be significant when P value is < 0•05.
When the levels of anaphylatoxins namely C3a, C4a and C5a proteins and Mannose Binding Lectin protein of complement system were analyzed in the serum of both the subjects and healthy individuals, it was found that only C5a and MBL titer was significantly higher in patients’ serum (Table 1,2,3,4 and figs 1,2,3 and 4). But the level of other anaphylatoxins namely C3a and C4a was not significant between the RHD subjects and the controls.
This was performed to qualitatively measure the presence of C3c and C3d in the serum samples of both the subjects and the controls. [Fig-5] and [Table-5] show the presence of C3c and C3d proteins in these two groups of samples. From the table it is clear that most of the patients were positive for C3c and C3d. None of the control samples were positive for the above products of C3b.
The complement system is one of the main effector mechanisms of innate immunity. As a key mediator of inflammation, its excessive or inappropriate activation through the classical, alternative or lectin pathways may contribute greatly to host tissue damage [3 & 5].
Ferreira & Nussenzweig [6] (1976) have described that C3 fulfills multiple physiological functions in host defense against pathogenic microorganisms and probably transformed host cells. It occurs in plasma and in other body fluids in inactive, but activatable form. The activation peptide, C3a, constitutes one of the known anaphylatoxins.
In addition to C3a, C3b also, have been implicated as mediators in the stimulation of antibody-forming cells [7 & 8]. The functions of C3a and C3b are controlled, respectively, by the serum enzymes carboxypeptidase B [9] (or anaphylatoxin inactivator) and C3b-inactivator [9 & 10]. Serum carboxypeptidase B removes the COOH-terminal arginyl residue from C3a [11] and C3b-inactivator cleaves C3b into the fragments C3c and C3d [12 & 13]. The biological significance of C3 in man has been documented by the observation of recurrent severe bacterial infections in a patient with inherited homozygous C3 deficiency [14] and in patients with congenital C3 hyper catabolism [15] due to a genetic deficiency of the C3b-inactivator [16] . More extensive proteolysis of C3b gives rise to the fragments C3c and C3d.
Ahmed et al., [17] (1990) proposed that C4a, an anaphylatoxin like C3a, is inactivated in vivo to C4a (des Arg) by serum carboxypeptidase N. Several methods have been developed to measure the split products of C3. The anaphylatoxin C3a, a split product of C3 generated during complement activation via the classical or alternative pathways, is inactivated rapidly by the removal of its N-terminal arginine by serum carboxypeptidase N, forming C3a (des Arg) [18] .
Like C3a, C5a can be used as an indicator of terminal sequence activation. C5a is rapidly (t1/2, 20 min) cleared from the circulation by C5a receptors on neutrophils even before inactivation by carboxypeptidase N. Thus, C3a measurement has advantages over C5a measurement in assessing terminal pathway activation.
The main role of the above anaphylatoxins is to induce anaphylactic reaction and hence we wanted to study their role in rheumatic heart disease. From our serum analysis of the anaphylatoxins, it was evident that C5a rather than C3a and C4a were observed significantly higher in subjects when compared to the normal individuals.
These findings suggest the presence of ongoing inflammation in these patients with probable complement activation due to the presence of more severe mitral stenosis or of an artificial valve.
Serum levels of C3d and immune complexes were shown to be increased in both the patients with acute RF (Rheumatic fever) and chronic RHD when compared to normal controls in previous study [19] . The most severe sequela of acute RF is chronic RHD. Previous reports have shown the deposition of complement proteins in heart tissue of patients with RHD is the leading cause of damage to the tissue [20] . In our study too, the presence of C3c and C3d in the patients and not in the normal individuals, explains that these proteins may also be one of the causative factors of damage to the heart tissue.
Guilherme L et al., [21] (2010) described that MBL is an acute phase inflammatory protein and functions as a soluble pathogen recognition receptor. MBL deficiency increases the overall susceptibility of individuals to conditions such as infectious diseases [22] , atherosclerosis [23] , leukemia [24] and abortion [25] . On the other hand, a high concentration of circulating MBL has been associated with infection by intracellular pathogens such as Leishmania and M. leprae [26 & 27]. Evidence has shown that MBL also play a role in autoimmune diseases such as systemic lupus erythematosus (SLE) [28] , ulcerative colitis [29] , Crohn’s disease [30] , Rheumatoid arthritis [31] and Sj¨ogren’s syndrome [32] .
In our study, this protein showed a significantly higher level when compared to that of normal individuals suggesting that MBL may represent a pathogenic factor in the complex physiopathology of the disease. Similar result was observed by Schafranski et al. [33] (2004) also.
The elevated MBL levels observed in patients with chronic RHD might corroborate the chronic inflammatory activity present in these individuals and contribute to valve injury through complement activation. MBL along with the other complement proteins may contribute for the complex physiopathology of the disease.
From the current study, it is evident that the complement proteins C5a, MBL, C3c and C3d may be the initiation factors of the disease since they are observed significantly more in the subjects and also contribute to the tissue damage. Since all the above analysis was done in samples of subjects who did not undergo treatment for RHD, the presence of the above proteins in the serum can be used as early markers of the disease and these patients can be protected from further damage of the heart tissue if they are diagnosed in the earlier stage itself and start the treatment as early as possible. Like other previous studies, our work also confirms the involvement of the complement proteins in tissue damage.
The authors acknowledge the patients and the healthy volunteers who gave their blood sample for the above analysis.
None
[1] Stollerman G.H. (1997) Lancet, 349, 935-942.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[2] Guilherme L. and Kalil J. (2008) Future Rheumatol, 3, 161-167.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[3] Walport M.J. (2001) N Engl J Med, 344, 1058-1066.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[4] Walport M.J. (1993) Br. J Rheumatol, 32, 269-273.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[5] Kirschfink M. (1997) Immunopharmacol, 38, 51-62.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[6] Ferreira A. and Nussenzweig V. (1976) Nature, 260, 613 – 615.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[7] Dukor P. and Hartmann K.U. (1973) Cell Immunol, 7, 349-356.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[8] Dukor P., Schumann G., Gisler R.H., Konig M., Hadding U. and BitterSuermann D. (1974) J. Exp. Med, 139, 337-354.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[9] Bokisch V.A., Muller-Eberhard H. and Cochrane C.G. (1969) J. Exp. Med. 129, 1109-1130.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[10] Tamura N. and Nelson Jr. (1967) J. Immunol, 99, 582- 589.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[11] Budzko D.B., Bokisch V.A. and Mfiller-Eberhard H.J. (1971) Biochem, 10, 1166-1172.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[12] West C., Davis N.C., Forristal J., Herbert J. and Spitzer R. (1966) J. Immunol, 96, 650-658.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[13] Ruddy S. and Austen K.F. (1971) J. Immunol, 107, 742-750.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[14] Alper C.A., Colten H., Rosen F.S., Rabson A.R., Macnab G.M. and Gear J.S.S. (1972) Lancet, 2, 1179-1181.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[15] Alper C.A., Abramson N., Johnston R.B., Jandl J.H. and Rosen F.S. (1970) N. Engl. J. Med, 282, 349-354.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[16] Abramson N., Alper C.A., Lachmann P.J., Rosen F.S. and Jandl J.H. (1971) J. Immunol, 107, 19-27.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[17] Ahmed A.E.E., Veitch J. and Whaley K. (1990) Immunol, 70, 139-144.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[18] Kohl J. and Bitter-Suermann D. (1993) Complement in health and disease, 299-324.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[19] Reddy K.S., Narula J., Bhatia R., Shailendri K., Koicha M., Taneja V., Jhingan B., Pothineni R.B., Malaviya A.N. and Mehra N.K. (1990) Indian J Pediatr, 57, 693-700.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[20] Prehm S., Herrington C., Nickel V., Volker W., Briko N.I., Blinnikova, Schmiedel A. and Prehm P. (1995) J. Anat, 187, 271-277.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[21] Guilherme L. and Kalil J. (2010) J Clin Immunol. 30 (1), 17-23.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[22] Eisen D.P. and Michinton R.M. (2003) Clin. Infect. Dis, 37, 1496-1505.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[23] Rugonfalvi-kiss S., Endresz V., Madsen H.O., Burian K., Duba J., Prohaszka Z., Karadi I., Romics L., Gonczol E., Fust G. and Garred P. (2002) Circulation, 106, 1071-1076.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[24] Schmiegelow K., Garred P., Lausen B., Andreassen B., Petersen B.L. and Madsen H.O. (2002) Blood, 100, 3757-3760.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[25] Christiansen O.B., Kilpatrick D.C., Souter V., Varming K., Thiel S. and Jensenius J.C. (1999) Scand J Immunol, 49, 193-196.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[26] Ambrosio A.R. and Messias-Reason I.J. (2005) Parasite Immunol, 27, 333-340.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[27] Dornelles L.N., Pereira-Ferrari L. and Messias-Reason I. (2006) Clin. Exp. Immunol, 145, 463-468.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[28] Huang Y.F., Wang W., Han J.Y., Wu X.W., Zhang S.T., Liu C.J., Hu Q.G., Xiong P., Hamvas R.M., Wood N., Gong F.L. and Bittles A.H. (2003) Eur J Immunogenet, 30, 121-124.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[29] Rector A., Lemey P., Laffut W., Keyaerts E., Struyf F., Wollants E., Vermeire S., Rutgeerts P. and Van Ranst M. (2001) Genes Immun, 2, 323-328.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[30] Seibold F., Konrad A., Flogerzi B., Seibold-Chmid B., Ami S., Juliger S. and Kun F.J. (2004) Gastroenterol, 127, 1076-1084.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[31] Ip W.K., Lau Y.L., Chan S.Y., Mok C.C., Chan D., Tong K.K. and Lau C.S. (2000) Arthritis Rheum, 43, 1679-1687.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[32] Wang Z.Y., Morinobu A., Kanagawa S. and Kumagai B. (2001) Ann. Rheum. Dis, 60, 483-486.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[33] Schafranski M.D., Stier A., Nisihara R. and Messias-Reason I.J.T. (2004) Clin. Exp. Immunol, 138 (3), 521–525.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
 | Fig. 1- This graph shows the mean C3a titre in the serum of patients and controls analyzed by Sandwich ELISA. |
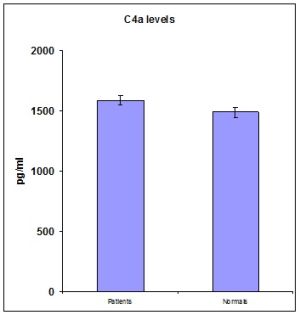 | Fig. 2- This graph shows the mean C4a titre in the serum of patients and controls measured by Sandwich ELISA. |
 | Fig. 3- This graph shows the mean C5a titre in the serum of patients and controls measured by Sandwich ELISA. |
 | Fig. 4- This graph shows the mean MBL level in the serum of patients and controls measured by Sandwich ELISA. |
 | Fig. 5- A representative Western Blot gel using serum from both the patients and controls hybridized with rabbit anti human C3c and rabbit anti human C3d antibodies. Lane 1-4 is the patients’ serum, Lane 5 is the control serum and Lane 6 is the protein marker. The patients’ serum samples were positive both for the C3c and C3d antibodies which is indicated by the 145kDa and 33kDa bands respectively. |
 | Table 1- C3a titre in the serum samples of both patients and controls |
 | Table 2- C4a titre in the serum samples of both patients and controls |
 | Table 3- C5a titre in the serum samples of both patients and controls |
 | Table 4- MBL titre in the serum samples of both patients and controls |
 | Table 5- The number of RHD samples positive for C3c and C3d in Western blot analysis. |