ISSN : 0975-5276
EISSN : 0975-9174

GUNASEKARAN P.1*, VANITHA M.C.2, MEERA B.3, RAMANI G.4, MALA RAO5, SEETA LAXMAN6
1Department of Genetics, Center for Excellence in Genomic Sciences, School of Biological Sciences, Madurai Kamaraj University, Madurai, India - 625021
2Department of Genetics, Center for Excellence in Genomic Sciences, School of Biological Sciences, Madurai Kamaraj University, Madurai, India - 625021
3Department of Genetics, Center for Excellence in Genomic Sciences, School of Biological Sciences, Madurai Kamaraj University, Madurai, India - 625021
4Department of Genetics, Center for Excellence in Genomic Sciences, School of Biological Sciences, Madurai Kamaraj University, Madurai, India - 625021
5National Chemical Laboratoires, Dr. Homi Bhabha Road, Pune - 411008
6National Chemical Laboratoires, Dr. Homi Bhabha Road, Pune - 411008
* Corresponding Author : gunagenomics@gmail.com
Received : 06-08-2011 Accepted : 02-09-2011 Published : 15-09-2011
Volume : 3 Issue : 2 Pages : 97 - 107
Int J Microbiol Res 3.2 (2011):97-107
DOI : http://dx.doi.org/10.9735/0975-5276.3.2.97-107
A cellobiohydrolase gene cbh1 was cloned from a cellulolytic fungus, Penicillum funiculosum NCL1. Nucleotide sequencing of cbh1 gene revealed that this gene was 1590 bp length encoding a putative protein consisting of 529 amino acids. The deduced amino acid sequence showed that the predicted molecular mass of the CBHI was 54.9 kDa, and showed significant homology to glycoside hydrolase family 7 cellobiohydrolases. The cbh1 gene was cloned using pET30b and expressed in E. coli BL21 (DE3). The expression analysis of the recombinant E. coli BL21 (pETC7) revealed the production of cbh1 transcript; however, functional cellobiohydrolase could not be detected. Therefore, the cbh1 gene was sub-cloned into GST tagged expression vector pGEX4t-3. The GST tagged cellobiohydrolase was purified to homogeneity using affinity chromatography. The recombinant enzyme exhibited optimum catalytic activity at pH 5.0 and 50 °C respectively. It was thermostable at 50 °C and retained 70% of its original activity after 30 min at 60 °C.
Penicillium funiculosum, cellulase, cellobiohydrolase, molecular cloning, affinity chromatography, molecular simulation
Plant biomass is considered as one of the most essential source of renewable carbon and it consists of cellulose (40-55%), hemicellulose (25-50 %) and lignin (10 - 40%) [39] . Among them, cellulose serves as the most abundant and renewable biopolymer on earth and its degradation by cellulases, followed by subsequent utilizations are important for global carbon sources [12] . The degradation of cellulosic materials has gained intense research and industrial interest due to its immense potential for converting cellulose to simple sugars [20] . Cellulolysis catalyzed by cellulases represents a major carbon flow from fixed carbon sinks to atmospheric carbon and it is highly warranted in the field of agricultural and wastewater treatment sectors and it can also be used to produce biobased products to replace the depleting fossil fuel [1,13] The microbial degradation of cellulose is achieved by cleaving of β-1, 4 - glycosidic bonds between the ß -D- glucose units by different enzymes. These enzymes have been grouped into 14 different families of glycoside hydrolases [5] . The cellulases are classified into endo and exoglucanases based on their mode of action. Initially, endoglucanases, (END) (E.C. 3.2.1.4) acts randomly and cleave the exposed positions of the cellulose chain in the amorphous regions to create new ends followed by the action of exoglucanases, also called as
cellobiohydrolases (E.C. 3.2.1.91), which degrade the polymeric chain of the cellulose either at the reducing or non reducing ends and produce cellobiose as final product. Finally, β- glucosidase completes the hydrolysis process by degrading cellobiose to glucose [42] . Endoglucanases and cellobiohydrolases can be easily distinguished by their substrate specificity. Endoglucanase shows high specificity towards soluble cellulose derivatives, such as CMC and very low specificity towards microcrystalline cellulose, whereas cellobiohydrolases (CBH) are important component of the multienzyme cellulase complex which show relatively higher specificity towards microcrystalline cellulose and responsible for the conversion of cellulose to soluble sugars. The synergistic action of both enzymes is required for complete conversion of cellulose to simple sugars [43] Glycoside hydrolases family 7 (GH7) contain both CBH and END types of enzymes. CBHI consists of a family GH7 catalytic module and a family GH1 carbohydrate-binding module (TrCBM1) connected by a proline- and threonine-rich, highly glycosylated linker peptide. The structure of CBH differs from END by loops of polypeptide chain covering the active site residue converting the active site cleft of the END into the characteristic tunnel of the CBH. The CBH belongs to three different glycoside hydrolase families namely GH6, GH7 and GH48. The importance of two cellobiohydrolases, CBH1 and CBHII has been realized for their preferences to act on the reducing and non-reducing ends of cellulose chains of microcrystalline cellulose [14,29,32] In fungi, the presence of CBH1 is needed for ordered degradation of cellulose, which requires all three structural modules of the cellulase i.e., the CD, C-terminal CBD, and linker region [33] .
The fungus Penicillium funiculosum is a filamentous fungus an efficient producer of cellulase [34] [27] . The P. funiculosum have been reported to produce high amount cellulases such as CMCase (13-15 U/ml), pNPGase (10-12 U/ml) and cellobiase (7.5 U/ml). Although improved strains of P. funiculosum have been isolated for overproduction of native enzymes, no genetically modified strains has been reported. Genes coding cellobiohydrolase GH7 have been cloned and characterized from a variety of fungal sources, including Humicola grisea [41] , Trichoderma reesei [38,35] , Penicillium janthinellum [21] , P. chrysosporium [6,9] and Aspergillus sp. (4,40). Here, we report the cloning and expression of cbh1 gene from P. funiculosum NCL1.
Penicillium funiculosum NCL1 was obtained from National Chemical Laboratory, Pune, India. Stock cultures were kept on potato dextrose agar and sub-cultured monthly. P. funiculosum NCL1 spores were inoculated in Reese Basal medium as described by [28] at a final concentration of 108 spores/ml. Flasks were incubated in an orbital shaker (220 rpm) at 30 ºC for 96 h. The mycelia were recovered by filtration on a nylon filter (30 μm spore) washed with 0.9% (w/v) NaCl and dried by pressing between two filter papers.
Restriction enzymes, T4 DNA ligase, Taq DNA polymerase were from MBI Fermentas (Opelstrasse, Germany). Bacterial strains and plasmids used in this study and their sources are listed in [Table-1] . E. coli DH5α (Invitrogen, CA, USA), E. coli BL21 (DE3) (Novagen, USA) were grown in Luria-Bertani (LB) broth or agar (Hi-Media). Whenever needed, the medium was supplemented with ampicillin (100 μg ml-1), Kanamycin (30 μg ml-1). The plasmids pTZ57R/T (MBI Fermentas, Opelstrasse, Germany), pET30b (Novagen) and pGEX4t-3 (Gift from Dr. Takashi Akiyama National Agricultural Research Center, Japan) were used in cloning and expression study. The primers used in this study are listed in [Table-2] .
Plasmid DNA was isolated from E. coli using a QIA prep spin plasmid preparation kit (Qiagen, Hilden, Germany) according to the manufacturer’s instruction. All restriction enzymes were purchased from MBI Fermentas (Opelstrasse, Germany). Agarose gel electrophoresis was performed as described by Sambrook et al (1989). DNA fragments from agarose gels were purified using the GEL extraction kit (Qiagen, Hilden, Germany).
PCR was performed in Mastercycler (Eppendorf, Germany) programmed for 30 cycles with one cycle of 94 °C for 5 min followed by 35 cycles of denaturation (60 s at 94 °C), annealing (60 s at 50 - 60 °C) and extension (60 s at 72 °C), with a final extension of 72 °C for 10 min. For analysis, 10 µl of reaction product was electrophoresed on 1% (w/v) agarose gel and stained with ethidium bromide (5 µg/ml).
Genomic DNA was isolated from the mycelia of P. funiculosum NCL1 using the method described by Murray and Thompson (1976). The DNA was resolved on agarose gel and was purified using gel extraction kit (Qiagen, Hilden, Germany) according to manufacturer’s instruction. The cbh1 gene was amplified from the genomic DNA using degenerate primers cbh7F and cbh7R, designed based on the conserved amino acid sequences of the corresponding GH families. The PCR product was then resolved on agarose gel and then purified. The purified PCR product was ligated in to pTZ57R/T and was used to transform E. coli DH5α. Both the strands of the cloned gene was completely sequenced using M13 primers (Macrogen, Seoul, Korea).
The cbh1 gene was PCR amplified without the leader sequence from the chromosomal DNA of P. funiculosum NCL1 using the forward primers pETcbh7F, pG7F and reverse primers pETcbh7R, pG7R. The cbh7 amplified product was digested with NdeI and XhoI restriction enzymes and ligated in to pET30b under the control of T7 promotor. Similarly, the pG7 amplified product was digested with BamHI and NotI and ligated with pGEX4t-3 digested with the same restriction enzymes. The recombinant plasmids were then used to transform E. coli BL21 (DE3) and the expression of cloned gene was induced with 0.4 mM of IPTG (Isopropyl-β-D-thio galacto pyranoside). The induced cells were harvested by centrifugation at 4 °C for 10 min at 12,000 X g and washed with 50 mM sodium acetate-buffer (pH 4.8). The cells were then disrupted by sonication (five times for 30 s with 30 s interval) (Labsonic U, Germany), and centrifuged at 12 000 X g for 30 min. The pellet was rinsed with 50 mM Sodium acetate buffer (pH 4.8) and stored at -20 °C until use. Both the soluble and pellet fractions were analyzed for CBH1 activity.
Cellobiohydrolase activity was measured by using avicel as substrate. One ml of the reaction mixture containing 0.5 ml of appropriately diluted enzyme and 0.5 ml of 1% avicel in 50 mM acetate buffer pH 4.8 was incubated at 55 °C for 30 min. After incubation, the reaction was terminated by addition of 1 ml DNS [30] . The absorbance was measured at 540 nm after dilution with 10 ml of distilled water. The reducing sugar released was estimated as glucose equivalents by DNS method. CBH activity was expressed as U/ml (μmol of reducing sugar formed per min) [28] . Protein concentration in the sample was measured by the Bradford method [3] .
The proteins were resolved on SDS-PAGE (12% v/v) according to Laemmli (1970). The gel was stained with Coomassie brilliant blue R-250 .The molecular mass of protein was determined in comparison with the mobility of the protein molecular weight markers (Amersham Biosciences, UK). For activity staining, the cellular proteins of recombinant strains were resolved on 12% polyacrylamide gel containing 100 µl of 50 mg/ml Methyl-Umbelliferyl-Cellobioside. After electrophoresis, the gel was immersed in 50 mM acetate buffer for 30 min and the gel was visualized under UV for detecting proteins exhibiting cellobiohydrolase activity.
Nucleotide and deduced amino acid sequences were analyzed with the Blast tools (www.ncbi.nlm.nih.gov/BLAST). Signal peptide sequence was analyzed using SignalP 3.0 [http://www.cbs.dtu.dk/services/SignalP] . Related sequences obtained from databases using the software GENSCAN online tool (www.genes.mit.edu/GENSCAN.html) were used for identification of gene features such as exon and splice sites in genomic DNA. BioEdit (version 7.0.4.1) was used for sequence editing and analysis. The structure of cbh7 was predicted using I-TASSER server [http//zhanglab.ccmb.med.umich.edu/I-TASSER] and the predicted structure was validated using protein structure validation software suite (PSVS) tool [2] . Molecular simulation of substrate Avicel with CBHI was carried out with the help of docking server using the Lamarckian genetic algorithm (www.dockingserver.com) .
Fresh LB kanamycin medium was inoculated with 1% of the overnight culture of recombinant E. coli BL21 and incubated at 37 ˚C until culture reaches 0.5 OD600nm. The culture was induced with different concentrations of IPTG (10 to 100 µM). After 6 h of induction, RNA was isolated. One step RT-PCR was performed to synthesis cDNA (Invitrogen, USA). The cbh1 gene was amplified from the cDNA using gene specific primers.
The recombinant E. coli BL21 was grown in LB medium for 24 h. After incubation, the cells were collected by centrifugation (5,000 X g for 15 min at 4°C) and supernatant were concentrated by ammonium sulphate precipitation and dialyzed. The recombinant cellobiohydrolase was purified using Fast Protein Liquid Chromatography system (Amersham Pharmacia Biotech, Piscataway, NJ). The purification of GST-tagged protein from the culture supernatant was performed using packed with Glutathione Sepharose GST-trap column (GE Healthcare Biosciences, USA) affinity chromatography. The culture supernatant was loaded on to 5 ml GST Hi-trap column equilibrated with 10 column volume of binding buffer (PBS; 140 mM NaCl, 2.7 mM KCl, 10 mM Na2HPO4, 1.8 mM K2HPO4, pH 7.3). The column was washed with the same binding buffer at a flow rate of 0.5 ml/min for 30 min to remove the unbound proteins and subsequently bound protein was eluted with elution buffer containing (50 mM Tris-HCl, 10 mM reduced glutathione, pH 8.0) at a flow rate of 1 ml/min. To cleave the GST-tag from the recombinant protein thrombin (a serine protease) was used. The purified recombinant protein was digested with thrombin for 12 h at 22°C to remove the GST region and then digested recombinant protein was loaded on to Hi-trap Benzamidine Fast Flow column chromatography. Fractions containing CBH activity were pooled and concentrated using Speed Vac concentrator (Thermo Scientific, Northridge, USA). The concentrate was dialyzed against 50 mM sodium acetate buffer (pH 4.8) for 4 h. Molecular mass and purity of the enzyme was confirmed by SDS-PAGE analysis.
The effect of pH on the CBHI activity was determined by measuring the relative activity at different pH with sodium citrate buffer (100 mM, pH 3-4.5), sodium acetate (100 mM, pH 4.5-5.5), and sodium phosphate (100 mM, pH 5.5-8.0)and sodium carbonate (8.0 -9.0). The effect of temperature on the reaction rate was determined by performing the standard reaction at different temperature from 30 to 90 ºC. The maximum activity was considered as 100%, and used as reference in determining relative activities.
The stability as a function of pH was determined by measuring their residual activity after 1 h of incubation at 37 ºC at each pH. Thermostability of CBHI was determined by incubating the enzymes at different temperatures (30 - 90 ºC) for 30 min. The residual activity was assayed at optimal pH 5 and temperature 50 ºC.
The substrate specificity of CBHI was determined by using, p-nitrophenyl-β-D-glucopyranoside (pNPG) and pNP-β-D-cellobiose (pNPC). CBH activities on polysaccharides such as, carboxy methyl cellulose (CMC), xylan, Avicel, Whatman filter paper were also tested. The p-nitrophenol released was determined under standard assay conditions. The activity on polysaccharides was estimated by assaying the amount of released glucose using the GOD-POD method [26] . The enzyme activity on polysaccharides was estimated by measuring released reducing sugars by the DNS method using glucose as standard.
The effects of various metal ions (BaCl2, MgCl2, MnCl2, ZnCl2, CaCl2, NiCl2, or CoCl2) and reagents (2-mercaptoethanol and cysteine) at 1 mM on CBH activity were determined by preincubating the enzyme with the reagents in 20 mM sodium acetate buffer (pH 5.0) at 30°C for 30 min. Activity was then measured at 50 °C for 15 min in the presence of the metal ions or reagents. The activity assayed in the absence of metal ions or reagents was recorded as 100%.
The Michaelis constant (Km) and the maximum velocity (Vmax) of CBHI were determined by carrying out the reactions with Avicel (0.5 to 50 mM) in 100 mM sodium acetate buffer (pH 5) at 50°C. Km and Vmax values were determined from Lineweaver-Burk plots using standard linear regression analysis.
Using P. funiculosum NCL1 genomic DNA as template, partial sequence of cbh gene was amplified with Cbh7F and Cbh7R primers. An expected amplicon of cbh was eluted from the gel and cloned in to pTZ57R/T vector. Both the strands of the insert in this plasmid were sequenced. The sequence exhibited homology towards the sequences of other fungal GH7 cellobiohydrolase in the GenBank and EMBL databases. To amplify the DNA fragments upstream and downstream of GH7 gene of P. funiculosum, were designed based on the sequence of GH7 partial gene and PCR amplified using the primers ORFcbh7F & ORFcbh7R and cloned into pTZ57R⁄T. Nucleotide sequencing of cbh gene revealed that this gene was 1590 bp length encoding a putative protein consisting of 529 amino acids. This CBH had an estimated molecular mass of 54.9 kDa and had signal sequence with 23 amino acids as predicted by the signal P. Multiple sequence alignment of this CBH with other known cellobiohydrolase sequences showed 99% identity with CBHI of P. occitanis [Fig-1] . At the N terminus, P. funiculosum CBHI consist of catalytic domain, the serine -theronine rich “hinge domain from Ser444 to Ser490 and cellulose binding domain from Gly491 to Leu530. The catalytic domain (50 - 60% identity) and the cellulose binding domain (35-70% identity) were well conserved among the CBHI, which showed that the P. funiculosum CBHI belongs to family GH7.
The cbh1 gene without signal sequence was amplified and cloned in to expression vector pET30b and the resultant recombinant plasmid was designated as pETC7. The cbh1 gene was expressed using T7 promotor in the recombinant E. coli BL21 (pETC7). Induction with different concentration of IPTG showed no new protein in either intracellular or extracellular fraction of the recombinant strain. However, transcript analysis revealed the expression of cbh1 gene specific transcript upon induction. Therefore, cbh1 was sub-cloned in another vector pGEX4t-3 and the resultant recombinant plasmid was transferred to E. coli BL21 (DE3). The cloning of the cbh1 in-frame with the GST tag allowed the formation of cloned gene product as GST fusion protein. The expression of the cbh1 gene was inducible with IPTG as this gene was cloned downstream to the tac promoter. Upon induction, the E. coli BL21 (DE3) harboring the recombinant plasmid expressed the cbh1 gene as a fusion protein with the glutathione-S-transferase (80.9 kDa). The intracellular fraction of the recombinant strain showed 1.6 U/ml of CBHI activity. Further, resolving proteins of the cellular fraction on SDS-PAGE, a new protein appeared corresponding to the expected size of 80.9 kDa with the GST tag. The recombinant protein from E. coli BL21 (DE3) was then purified using GST trap column and was resolved on SDS-PAGE [Fig-3] . The purified protein exhibited maximum activity of 3.57 U/mg. The recombinant CBHI showed activity towards the MUC in zymogram.
The effect of different pHs and temperature on CBHI activity was determined with Avicel as substrate. The purified CBHI was incubated at 50 °C for 1 h at different pH (3-10). The results showed the optimum pH for the CBHI was 5.0, with 95% and 90% of the maximum activity appearing at pH 4.5 and 5.5 respectively. An acidic pH optimum and maximal activity at about pH 4.5 are common features of similar CBH from diverse microbial organism. The isoelectric point pI, was determined to be 4.62, which is typical for extracellular CBHs. The optimum temperature for the hydrolysis reaction was 50 °C while 75% and 95% of the maximum activity was found at 45 and 55°C respectively [Fig-4] .
The activities of P. funiculosum CBHI with various substrates are shown in [Table-3] . This CBHI showed maximum activity on pNPC (pNP-β-D-cellobioside) but showed only 0.2%, 62.2% and 75.8% activity on pNPG, Avicel, and filter paper respectively. Km value of CBHI of P. funiculosum was found to be 0.9 mM pNPC. CBHI did not show any activity on xylan.
The effects of metal ions and other additives on the CBHI activity were examined and the results are presented in [Table-4] . Among several metal ions examined, only Ca2+ stimulated the CBHI activity significantly. MnCl2, MgCl2, NiCl2, BaCl2, ZnCl2, CaCl2, or CoCl2 did not affect the CBHI activity.
Homology modeling was carried out using SWISS-MODEL software and T. emersonii cel7 (IQ9H) as a template to predict the structural characteristics of P. funiculosum CBHI. The Ramachandran plot showed that 90.9% (479/527) of all residues were in favoured (98%) regions. The quality of the predicted structure of CBHI was validated using the protein structure validation software suite (PSVS). Ramachandran plot obtained for CBHI indicated that 90.1% of the amino acids had favourable regions [Fig-2] . A, B and L indicate the most favoured amino acid conformations (81.1%), while a, b and l indicate unfavourable conformations. ~a, ~b, ~l and ~p denote generously allowed amino acid conformations (1.3%). Molecular simulation of the CBHI with substrates Avicel and CMC using docking studies revealed that, the CBHI was able interact with 12 different amino acids i.e ASN: 62, TRP: 63, ARG: 129, TYR:167, ASP: 195, GLN: 197, PRO: 199, ASP: 201, GLU: 234, TYR: 269, SER: 394, TRP: 396 for the substrate Avicel and 8 different amino acid residues i.e., TRP: 143, PRO: 230, ASP: 231, ASP: 273, ALA: 314, ASN: 315, TRP: 377, LYS: 405 for the substrate CMC with an electrostatic energy of -0.15 kcal/mol for Avicel and -0.48 kcal/mol for CMC and intermolecular energy of-6.28 kcal/mol for avicel and -4.95 kcal/mol for CMC with an electrostatic energy of -0.15 kcal/mol and intermolecular energy of -6.28 kcal/mol.
The nucleotide sequence of the cbh1 gene of P. funiculosum NCL1 was deposited in the GenBank database under the accession number HQ615690.
The cellobiohydrolases (CBH) of family GH-7 are found exclusively in eukaryotic organisms. Of these, filamentous fungi seem to be the major producer of this class of enzymes. The purification and properties of CBH from several filamentous fungi have been reported but there are no reports on the genes encoding CBH from P. funiculosum. Therefore, cbh gene of P. funiculosum was cloned and expressed; further, the gene product was purified and characterized. The molecular mass of CBHI from P. chrysogenum FS010 and Penicillium occitanis was approximately 50 kDa [7,45] . The deduced amino acid sequence of P. funiculosum CBHI to those of other fungal CBHIs showed around 99% identity with CBHI of P. occitanis and 98% with XynA, xylanase/cellobiohydrolase of P. funiculosum. The multiple sequence alignment of deduced amino acid sequence of P. funiculosum CBHI with other fungal CBH revealed that P. funiculosum consist of catalytic domain, the serine-theronine rich “hinge domain from Ser444 to Ser490 and cellulose binding domain from Gly491 to leu530 .The catalytic domain (50-60% identity) and the cellulose binding domain (35-70% identity) was well conserved among the CBHI, which showed that, the P. funiculosum CBHI belongs to GH7 [15] . The 3D structure of the C terminal domain of CBHI from T. reesei was determined by NMR spectroscopy, which revealed the amino acid residues essential for cellulose-CBM interaction. The three aromatic residues (Tyr524, Tyr525, Tyr498) within the cellulose binding domain and the side chains of the three amino acids (Gln500, Asn522, Gln527) which stabilizes the cellulose-CBM interaction conserved among the CBHI cellulases were also found to be present in P. funiculosum NCL1. A deduced signal peptide was found in the N-terminus from (Met to Gln27). The family GH7 enzymes were characterized by 22 conserved cysteine residues and two glutamic acid residues in their catalytic site which were also found to be conserved in P. funiculosum NCL1, suggesting the enzyme as CBHI type.
To elucidate the structure of CBHI and to study their interactions with Avicel, 3-D model of CBHI was constructed on the basis of the X-ray crystallographic structure of cellobiohydrolase from T. emersonii (1Q9H) which shares 72% identity with P. funiculosum. Ramachandran plot of CBHI showed that, 90.9% (479/527) of all residues of CBH1 were in favoured (98%) regions suggesting the higher stability of the predicted structure. 1Q9H is a single-domain protein consists of β-sandwich structure the convex and concave sheets of the β-sandwich are composed of seven β-strands and the loops extending from the β-sandwich forms a tunnel, which runs the length of the concave sheet, into which the cellulose substrate can be accommodated, the β sandwich represents the characteristics of Family GH7 [10] . The recombinant E. coli BL 21 (pETC7) induced with different concentration of IPTG did not over produce the CBHI. However, transcript analysis of the same recombinant strain revealed the expression of cbh1 gene specific transcript. Similar results were observed with respect to the expression of cbh1 in E. coli [15] . Few other eukaryotic proteins were expressed in E. coli and accumulated within the cells as insoluble aggregates [11] . The CBH specific cDNA from P. chrysosporium was cloned and expressed in E. coli using pET cloning and expression system, which required refolding of inactive protein product by in vitro chemical methods to restore its enzymatic activity [17] . In order to eliminate the refolding problems, CBHI was heterologously expressed using glutathione S-transferase (GST) fusion system [18] . Similarly cbh1 was sub-cloned into GST tagged vector pGEX4t-3 and expressed in E. coli BL21 (DE3). The GST tagged protein facilitated easy purification of the recombinant protein from E. coli BL21 (DE3) using GST trap column. The purified protein exhibited molecular weight of 54 kDa without GST tag in SDS-PAGE with maximum activity of 4.2 U/ml [44] . Similarly, the β-1,3-1,4 glucanase has been also purified as fusion protein using GST trap FF coloumn. The CBHI enzyme showed maximum activity at pH 5. The optimum pH reported for other fungal CBH range from 4.0 to 6.0 [18] . The optimal temperature for activity was observed at 50oC with 99% and 22% of the maximum activity at 50 and 60°C respectively. In T. versicolor, the purified CBHI had an optimum pH and temperature of 5.0 and 40 ºC respectively. The enzyme was stable over a wide pH range extending from 3.0 to 9.0 and at temperatures lower than 50o C [19] It has been reported that F. pinicola CBH had an optimum temperature 55oC and pH 5 [37] . The pH optima of rumen fungal cellulases range from 5.5 to 6.5 and temperature from 40 to 50 oC [25] . The relative rate of hydrolysis of the purified CBHI from P. funiculosum NCL1 against various substrate showed that, the highest activity was observed with pNPC, followed by avicel. In P. purpurogenum, the purified CBH showed broad substrate specificity with maximum activity towards p-nitrophenyl β-D-cellobiopyranoside [22] . The enzyme activity of purified CBHI was measured in the presence of metal ions or other additives, which showed that Ca2+ efficiently stimulates the enzyme activity than other metal ions. Similarly the F. pinicola cellobiohydrolase activity was not stimulated by BaCl2, MgCl2, MnCl2, ZnCl2,CaCl2, NiCl2, or CoCl2, and it was neither inhibited nor activated by EDTA [37] . The Km values of purified CBHs for pNPC from other fungi range from 0.58 to 6.8 Km value while the same of P. funiculosum NCL1 was slightly higher value.
Authors thank Department of Biotechnology, New Delhi, India for the financial support through a grant (No5/258/51/2006 -NMITLI). Author thanks UGC, India for the research fellowship under the scheme for meritorious students in Biosciences (F.No.4- 1/2006(BSR)/5-67/2007). The Centre for Advanced studies in Functional Genomics, Centre for Excellence in Genomic Sciences and Networking Resource Centre in Biological Sciences are gratefully acknowledged for support facilities.
[1] Berner R.A. (2003) Nature, 426, 323–6.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[2] Bhattacharya A., Tejero R. and Montelione G.T. (2007) Proteins, 66, 778-795.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[3] Bradford M.M. (1976) Analytical Biochemistry, 72, 248–254.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[4] Chikamatsu G., Shirai K., Kato M., Kobayashi T. and Tsukagoshi N. (1999) FEMS Microbiology Letters, 179, 239–245.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[5] Coutinho J.B., Gilkes N.R., Kilburn D.G., Warren R.A.J. and Miller R.C. Jr.(1993) FEMS Microbiology Letters, 113(2) , 211–217.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[6] Covert S.F., Bolduc J. and Cullen D. (1992) Current Genetics, 22, 407-413.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[7] Fatma B., Ali G., Mamdouh B.A., Hafedh B., Monia B. and Semia E.C. (2010) Enzyme Microbial Technology, 46 ,74-81.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[8] Gielkens M.M., Dekkers E., Visser J. and De Graaff L.H. (1999) Applied Environmental Microbiology, 65, 4340– 4345.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[9] Grassick A., Murray P.G., Thompson R., Collins C.M., Byrnes L., Birrane G., Higgins T.M. and Tuohy M.G.(2004) European Journal of Biochemistry, 271, 4495-4506.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[10] Guise A.D., West S.M. and Chaudhuri J.B. (1996) Molecular Biotechnology, 6, 53–64.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[11] Guo R., Ding M., Zhang S.L., Xu G.J. and Zhao F.K. (2008) Journal of Comparative Physiology, 178, 209-215.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[12] Haight M. (2005) Water Science Technology, 52, 553–9.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[13] Henrissat B., Driguez H., Viet C. and Schulein M. (1985) Biotechnology, 3, 722-726.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[14] Henrissat B. and Bairoch A. (1993) Biochemistry journal, 293, 781-788.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[15] Howard R.L., Masoko P., Mowa M.B, Abotsi E. and Howard S. (2004) African Journal of Biotechnololgy 3(7) , 349-352
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[16] Howard R.L. (1997) Cloning and expression of Phanerochaete chrysosporium cbhI-like cDNAs in Escherichia coli. Ph.D thesis, University of Manchester, United kingdom.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[17] Howard R.L., Masoko P., Mowa M.B., Abotsi E. and Howard S. (2004) African Journal Biotechnology, 4, 1180-1184.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[18] Karim L., Reginald S., Zhizhuang X., Kwang B.J., Yun Z., Justin P., Adrian T. and Luc V. (2007) Applied Microbiology and Biotechnology, 75(2), 337-346.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[19] Kim K.C., Seung S.Y., Oh Y.A. and Seong J.K. (2003) Journal of Microbiology. Biotechnology, 13, 1-8.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[20] Koch A., Weigel C.T.O and Schulz G. (1993) Gene, 124, 57-65.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[21] Lee K.M., Joo A.R., Marimuthu J., Lee K.M., Moon H.J. and Lee J.K. (2011) Applied Microbiology and Biotechnology 163, 25-39.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[22] Laemmili I.K. (1970) Nature, 227,680-685.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[23] Li Y.L., Li D.C. and Teng F.C. (2006) Wei Sheng Wu Xue Bao, 46: 143-146.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[24] Li X.L., Chen H. and. Ljungdahl L.G. (1997) Applied Environmental Microbiology 63, 4721–4728.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[25] Lin J., Balakrishna P. and Suren S. (1999) Biotechnology and Applied Biochemistry, 30, 81-87.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[26] Mala R. and Chittra M. (1984) Biotechnology Letters, 6(5), 319-332.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[27] Mandel M. and Reese E.T. (1960) Journal of Bacteriology, 79, 816.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[28] Medve J.J. Ståhlberg J. and Tjerneld F. (1994) Biotechnology Bioengineering, 44, 1064–1073.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[29] Miller G.L. (1959) Analytical Chemistry 31, 426-428.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[30] Murray M.G. and Thompson W.F. (1980) Nucleic Acids Research, 8, 4321-4325.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[31] Nidetzky B., Zachariae W., Gercken G., Hayn M. and Steiner W. (1994b) Enzyme Microbial Technology, 16, 43–52.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[32] Rabinovich M.L., Meinik M.S. and Bolobova A.V. (2002) Applied Biochemistry and Microbiology, 38, 305-321.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[33] Sahasrabudhe N.A., Lachke A.H. and Ranjekar P.K. (1985) Biotechnology Letters 9, 881-886.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[34] Saloheimo M., Lehtovaara P., Penttila M., Teeri T.T, Stahlberg J., Johansson G., Pettersson G., Claeyssens M., Tomme P. and Knowles J.K (1988) Gene, 63, 11–21.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[35] Sambrook J., Fritsch E.F. and Maniatis T. (1989) 2nd edn. Cold Spring Harbor, N.Y; Cold Spring Harbor Laboratory Press.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[36] Shin K., Yoon H.K., Marimuthu J., Jung-K.L. and Yeong S.K. (2010) Journal of Microbiology and Biotechnology, 20, 1681–1688.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[37] Shoemaker S., Schweickart V., Ladner M., Gelfand D., Kwok S., Myambo K. and Innis M. (1983) BioTechnology, 1, 691–696.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[38] Sun Y., Cheng J. (2002) Bioresource Technology, 83, 1-11.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[39] Takada G., Kawaguchi T., Sumitani S. and Arai M. (1998) Journal of Fermentation and Bioengineeering, 85, 1–9.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[40] Takashima S., Nakamura A., Hidaka M., Masaki H. and Uozumi T. (1999) Journal of Biochemistry, 125, 728–736.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[41] Wilson D.B., Irwin D.C. (1999) Advances in Biochemical Engineering Biotechnology, 65, 1-21.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[42] Yasokawa D., Takeshi S., Ryoji N., Takatugi I. and Koji N. (2003) Bioscience Biotechnology Biochemistry, 67, 1319-1326.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[43] Youn J.J., Yong S.L., In H.P., Subhosh C.M., Keun K.K. and Yong L.C. (2010) Indian journal of biochemistry & biophysics, 47, 203-210.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[44] Yunhua H., Tianhong W., Hao L. and Huiyuan Z. (2007) Acta Biochimica et Biophysica Sinica 39(2), 101–107.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
 | Fig. 1- Protein sequence alignment with ClustalW program of cellobiohydrolases from glycoside hydrolase family 7 Residues identical in all sequences are printed in white on a red background. Residues identical or with a conservative substitution in at least four of the seven sequences are printed in red on a white background. The Tyr side chains of the CBM (Y524, Y525) are shown by blue arrows and the side chains of three amino acids (Q500, N522, and Q527) are shown in brown arrow. The CBM and linker sequences are boxed. Penicillium funiculosum NCL1 CBHI exhibited an identity of 99%, 98%, 74%, 91% and 56% with Penicillium occitanis CBHI (AAT99321.1), Penicillium funiculosum xyn/cellobiohydrolase) (AJ312295), Talaromyces emersonii (pdb 1Q9H), Penicillium marneffi (XP_002149324.1), Talaromyces stipitatus (XP_002484839) respectively. |
 | Fig. 2- Molecular simulation of CBH7 with substrates Avicel and CMC Molecular simulation was carried out with the docking server using the Lamarckian genetic algorithm. |
 | Fig. 3- SDS-PAGE and Zymogram analysis of the purified cellobiohydrolase A) Affinity chromatography purified cellobiohydrolase was resolved on 12% SDS-PAGE and stained with Coomassie brilliant blue R-250. Lane 1- purified cellobiohydrolase (54.9 kDa). B) Zymogram analysis was performed on MUC incorporated separating gel. Lane 2- Zymogram analysis of purified cellobiohydrolase using MUC as substrate. Lane M- Molecular weight markers (from the top): β-galactosidase (116 kDa), Bovine serum albumin (66.0 kDa), ovalbumin (45.0 kDa), lactate dehydrogenase (35.0 kDa), REase Bsp981 (25.0 kDa), β-lactoglobulin (18.0 kDa) and lysozyme (14.0 kDa). |
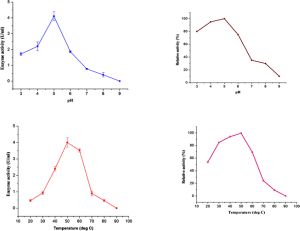 | Fig. 4- Effect of pH and temperature on the activity of cellobiohydrolase Cellobiohydrolase activity (A) was determined in different buffers (pH 3.0 - 9.0) at 50 °C. The stability of the enzyme at different pH levels (A) was determined by incubating the enzyme in different buffers for 1 h at 50 °C and the residual activity was measured at pH 4.8; 50 °C (C). The Cellobiohydrolase activity was measured at different temperatures (30 - 90 °C) (B) at pH 4.8. The purified enzyme was preincubated at different temperatures for 30 min and the residual activity was measured at pH 4.8 and 50 °C (D) Relative activity was calculated by considering the maximum activity as 100%. |
 | Table 1- Bacterial strains and plasmids used in this study |
 | Table 2- List of primers used in this study |
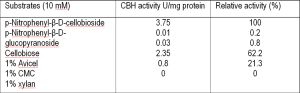 | Table 3- Substrate specificity of purified cellobiohydrolase from P. funiculosum NCL1 |
 | Table 4- Effect of metal inhibitors on purified cellobiohydrolase from P. funiculosum NCL1 |